
【概要】
炭素は生命体を構成する重要な元素の一つであり、植物は光合成により空気中の二酸化炭素から糖を合成し利用しています。しかし多くの生物が炭素を獲得するには、植物や他の生物から有機化合物を摂取する必要があります。細菌や菌類などの病原微生物が感染し増殖する際にも同様で、特に植物に感染する病原体は植物が光合成産物として高濃度に蓄積した糖を主な炭素源として吸収し利用しています。しかし病原体が植物から糖を搾取する一方、植物がそれに対抗する防御手段を備えているかはこれまで明らかとされていませんでした。
今回、京都大学大学院農学研究科の高野義孝教授、徳島大学大学院生物産業学研究部の山田晃嗣特任助教(元・京都大学大学院農学研究科 日本学術振興会特別研究員PD)、奈良先端科学技術大学院大学バイオサイエンス研究科・JSTさきがけの西條雄介准教授、理化学研究所環境資源科学研究センターの中神弘史ユニットリーダーによる共同研究グループは、病原体が感染した際に植物細胞は糖吸収活性を増強させることで細胞外の糖を回収し、病原体の糖へのアクセスを阻害する機構を見出しました。本成果は米科学誌「Science」へ掲載されるのに先立ち、オンライン版(2016年11月24日付け)に掲載されます。
1.背景
植物は光合成により空気中の二酸化炭素から糖を合成し利用できる一方で、多くの生物は植物や他の生物から有機化合物を摂取して炭素を獲得する必要があります。特に細菌や菌類などの植物病原体が植物に感染した際には、光合成産物として蓄積している植物の糖を主な炭素源として吸収し利用しています。多くの植物病原細菌は葉に侵入後、植物細胞の隙間で増殖します。私たちは植物が細胞外空間の糖を細胞内へ回収することで病原細菌の糖摂取を阻害しているのではないかと考え、防御応答活性化時の植物細胞の糖の取り込み活性に着目して解析を行いました。
2.研究手法と成果
初めに私たちは、モデル植物シロイヌナズナを用いて植物の糖吸収が病原細菌への抵抗性と関与しているかを調べました。糖吸収は細胞の脂質二重膜に埋め込まれている糖トランスポーター(輸送体)タンパク質によって行われます。そしてシロイヌナズナの糖吸収はSTP1およびSTP13の二つの糖トランスポーターによって主に行われていることを研究チームはすでに明らかにし報告していました。今回そのSTP1およびSTP13糖トランスポーターによる糖吸収が病原細菌への抵抗性に関与するかを、stp1 stp13遺伝子破壊植物に病原細菌を接種し細菌抵抗性を測定することで検証しました。その結果stp1 stp13遺伝子破壊植物では野生型植物よりも細菌抵抗性が低下していたことから、糖吸収と細菌抵抗性の関連性が示されました。
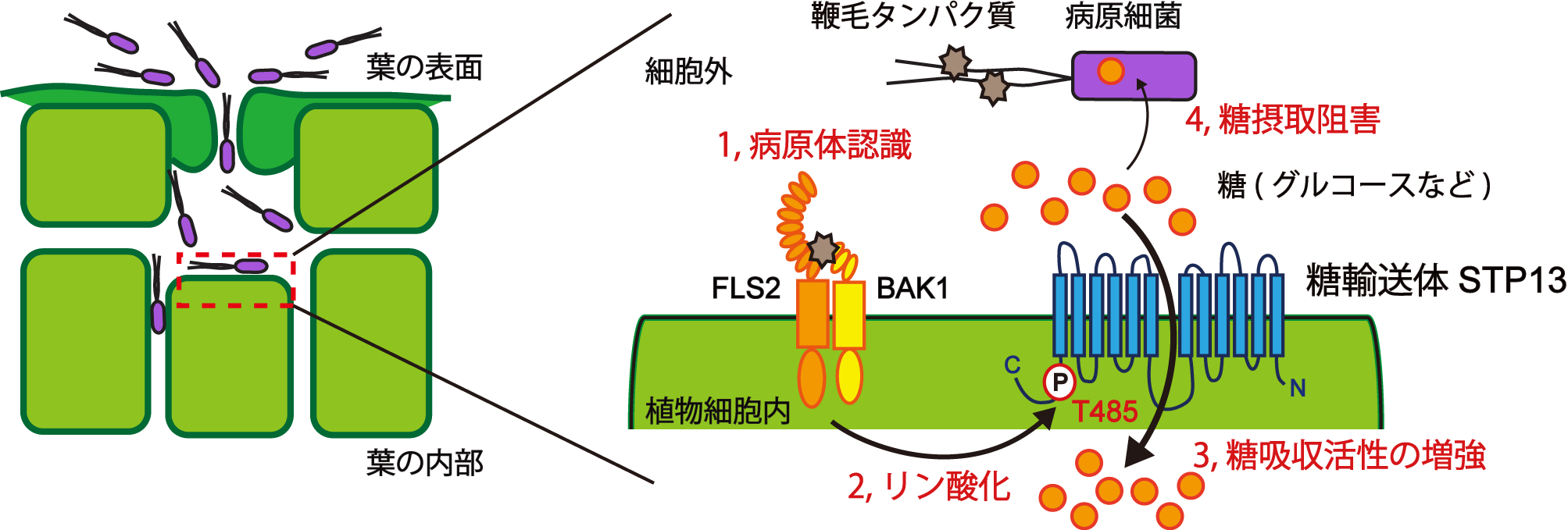
細菌の鞭毛タンパク質や菌類の細胞壁断片などの病原微生物に特徴的な分子を植物細胞が認識することにより防御応答は発動されます。私たちはシロイヌナズナが防御応答を活性化させた際に、既知の一連の防御反応に加えて糖吸収活性が著しく上昇することを見出しました。この結果は上で述べた、植物が病原細菌の侵入時に細胞外の糖を細胞内へ回収することで病原細菌の糖摂取を防ぐ、という考えを支持するものでした。さらに私たちは、この防御応答依存的な糖吸収にはSTP13糖トランスポーターが関与しており、STP13の糖吸収活性が防御応答の発動により制御されることを見出しました。
そして、このSTP13の制御機構について、まず私たちは細菌鞭毛タンパク質を認識するシロイヌナズナの受容体(FLS2)とその制御因子であるBAK1がSTP13と結合することを発見しました。続いて、BAK1はSTP13の485番目のアミノ酸であるスレオニン残基(T485)をリン酸化すること、さらにそのリン酸化はSTP13の糖吸収活性を増加させることを明らかとしました。STP13をstp1 stp13遺伝子破壊植物に導入すると細菌抵抗性が野生型植物並みに戻るにもかかわらず、T485がリン酸化されない変異型STP13をstp1 stp13遺伝子破壊植物に導入しても細菌抵抗性は戻りませんでした。この結果より、STP13のT485リン酸化による活性増強がシロイヌナズナの細菌抵抗性に重要であることが示されました。
次に、植物細胞外の糖含量の減少がどのように病原細菌の増殖の抑制につながるのかを調べました。病原細菌は病原性因子を植物細胞内に分泌することで植物の防御応答を抑制し感染を可能にしています。病原性因子の分泌は、病原性細菌が植物組織中に侵入した際に植物細胞の隙間に蓄積している糖を認識することで活性化します。私たちは病原細菌がstp1 stp13遺伝子破壊植物に感染する際、病原性因子の分泌が野生型植物に感染した時と比べて上昇していることを見出しました。stp1 stp13遺伝子破壊植物にSTP13を導入することで病原性因子の分泌上昇は抑えられましたが、変異型STP13の導入では抑えることができませんでした。そのため、リン酸化によるSTP13の糖吸収活性の増強を介して細胞外の糖含量を減少させることで病原細菌が糖にアクセスしづらくなり、病原性因子の分泌が抑制されていることがわかりました。また糖は病原性因子の分泌を活性化させる一方で、細菌の重要な代謝エネルギー源として利用されています。病原性因子を分泌できないように改変した病原細菌においてもstp1 stp13遺伝子破壊植物内では野生型植物内に比べて増殖が著しく増加していたため、stp1 stp13遺伝子破壊植物の組織内は細胞外に糖が豊富で病原細菌が増殖しやすい環境になっていることが考えられました。以上の結果より、今回私たちは、(i) 植物は防御応答の発動により糖トランスポーター活性を増強し細胞外の糖含量を減少させること、(ii) その結果として病原細菌の病原性因子の分泌抑制と代謝エネルギー制限につながり、病原細菌の増殖が抑えられることを明らかとしました。
3.波及効果、今後の予定
今回、細胞外部の糖を回収し病原細菌の糖吸収を阻害する植物の新規防御応答を明らかとしました。病原細菌に限らず多くの植物病原体は炭素源として糖を利用しているため、幅広い細菌および菌類に対してもこの防御が有効であることが考えられます。そのため本成果を基盤とし、植物の糖吸収を高める化合物を発見できれば、より広範囲の病原体に有効な新規農薬の開発につながる可能性があると期待されます。
4.研究プロジェクトについて
本成果は日本学術振興会(JSPS)科学研究費補助金、科学技術振興会機構(JST)戦略的創造研究推進事業個人型研究(さきがけ)および旭硝子財団助成金の支援を受けて行われました。
<論文タイトルと著者>
タイトル:Regulation of sugar transporter activity for antibacterial defense in Arabidopsis
著者:Kohji Yamada*, Yusuke Saijo, Hirofumi Nakagami, Yoshitaka Takano* (*,責任著者)
掲載誌:Science(http://science.sciencemag.org/content/early/2016/11/21/science.aah5692)
naistar:http://hdl.handle.net/10061/11184(NAIST Academic Repository:naistar)
<イメージ図>

<お問い合わせ先>
高野義孝・京都大学農学研究科・教授
TEL: 075-753-6131 FAX : 075-753-6131
E-mail: ytakano@kais.kyoto-u.ac.jp