
根が重力方向に曲がる新たな仕組みを解明
-インドール酪酸から作られるオーキシンの新たな機能の発見-
理化学研究所(理研)環境資源科学研究センター適応制御研究ユニットの瀬尾光範ユニットリーダー、渡邊俊介基礎科学特別研究員、奈良先端科学技術大学院大学の梅田正明教授、東京農工大学の笠原博幸教授、岡山理科大学の林謙一郎教授らの共同研究グループ※は、植物の根の重力屈性[1]に寄与するオーキシン[2]の微量な前駆体であるインドール酪酸(IBA)の細胞内取り込み輸送体[3](膜タンパク質)を発見しました。
本研究成果は、土壌から水分や栄養分を獲得する器官である根の重力応答機構の一端を明らかにし、植物の生産機能の向上などに貢献すると期待できます。
植物の根は重力を感じ取り、地中に向かって根を伸ばします。この現象は、植物ホルモン[4]のオーキシンが根端[5]の重力側に多く分布すること(不等分布)で誘導されます。植物体内における主要なオーキシンはインドール酢酸(IAA)であり、そのほとんどはインドールピルビン酸から合成されますが、ごく少量はIBAからも合成されます。
今回、共同研究グループは、IBAの細胞内取り込み輸送体として、シロイヌナズナ[6]の輸送体ファミリーNITRATE TRANSPORTER 1/PEPTIDE TRANSPORTER FAMILY (NPF)[7]の一つである「NPF7.3」を新たに同定し、IBAから合成されるIAAが重力屈性に重要な役割を果たしていることを明らかにしました。
本研究は、科学雑誌『Proceedings of the National Academy of Sciences of the United States of America (PNAS )』オンライン版(11月20日付)に掲載されました。
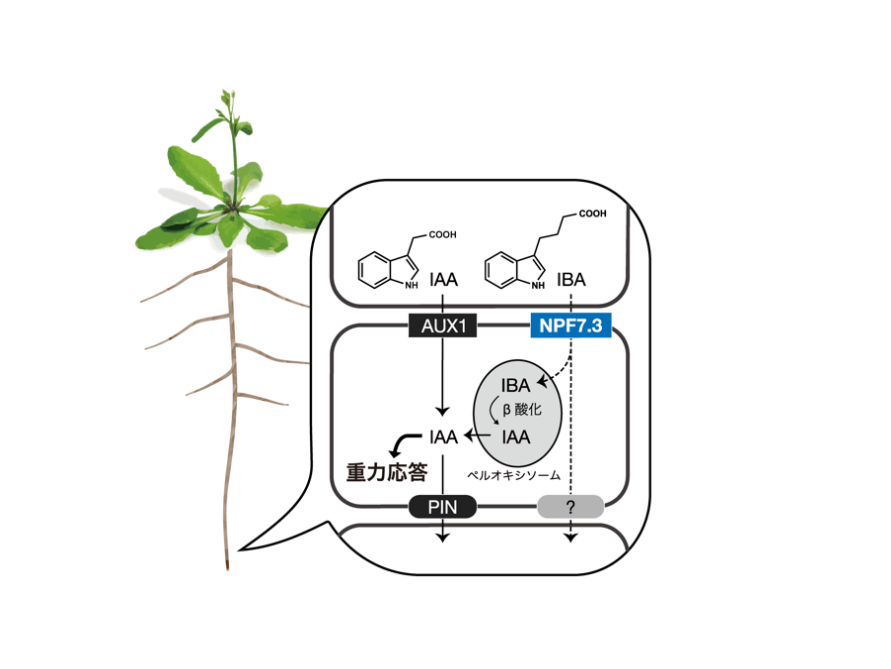
インドール酪酸(IBA)の輸送体NPF7.3による重力屈性の誘導機構仮説
※共同研究グループ
理化学研究所 環境資源科学研究センター
- 適応制御研究ユニット
ユニットリーダー 瀬尾 光範 (せお みつのり)
基礎科学特別研究員 渡邊 俊介 (わたなべ しゅんすけ)
特別研究員 鈴木 洋弥 (すずき ひろみ)
テクニカルスタッフⅡ 菅野 裕理 (かんの ゆり)
- 技術基盤部門 質量分析・顕微鏡解析ユニット
上級技師 豊岡 公徳 (とよおか きみのり)
テクニカルスタッフⅡ 武田 紀子 (たけだ のりこ)
- 奈良先端科学技術大学院大学 先端科学技術研究科
教授 梅田 正明 (うめだ まさあき)
助教 髙橋 直紀 (たかはし なおき)
- 東京農工大学 グローバルイノベーション研究院
教授 笠原 博幸 (かさはら ひろゆき)
大学院生 青井 勇輝 (あおい ゆうき)
- 岡山理科大学 理学部
教授 林 謙一郎 (はやし けんいちろう)
研究支援
本研究は、文部科学省新学術領域研究(研究領域提案型)「植物の生命力を支える多能性幹細胞の基盤原理(研究領域代表者:梅田正明)」の計画研究「多能性幹細胞の維持・再生機構の解明(研究代表者:梅田正明)」、日本学術振興会(JSPS)科学研究費補助金基盤研究(C)「ストレスに応答した植物の細胞周期停止を制御する分子基盤の解明(研究代表者:髙橋直紀)」などによる支援を受けて行われました。
1.背景
植物は成長に必要な水や無機栄養の大部分を、土壌から根を通して吸収しています。そのため、植物の成長には、環境に適した根系構造を発達させることが重要です。
重力は植物が受け取る環境刺激の一つであり、根は重力方向に沿って伸長します。この根の重力屈性には、植物ホルモンのオーキシンが重要な役割を果たすことが広く知られています。オーキシンが根端の重力側に多く蓄積すること(不等分布)が引き金となり、重力側の細胞伸長が抑制されることで、根は屈曲します。
1880年、ダーウィン父子が「植物の屈性を制御する何らかの信号」の存在を示唆して以来、オーキシンの発見を皮切りに、重力応答に関する情報伝達機構や代謝経路、輸送メカニズムが明らかにされてきました。特に、オーキシンの不等分布形成の要として、主要な天然オーキシンであるインドール酢酸(IAA)の輸送が長らく注目を浴びてきました。
植物体内で、IAAのほとんどはインドールピルビン酸から合成されますが、ごく少量はインドール酪酸(IBA)からも合成されます。IBAからIAAが合成される経路は主要ではありませんが、側根[8]形成などに特異的な働きを持つと考えられていました。しかし、この生合成経路が重力応答に果たす役割は分かっていませんでした。
2.研究手法と成果
共同研究グループは、複数の植物ホルモンの輸送に関わることが知られているシロイヌナズナの輸送体ファミリー「NITRATE TRANSPORTER 1/PEPTIDE TRANSPORTER FAMILY;NPF)」に着目しました。NPFは、生体膜を貫通し孔を形成する膜タンパク質で、膜内外の化合物の移動を仲介する働きをしています。まず、NPFの変異がシロイヌナズナの根の形態に与える影響を調べたところ、NPF の一つで細胞膜に存在する「NPF7.3」の機能を失ったnpf7.3変異体では、根が重力方向に沿って直線的に伸長しないこと、npf7.3変異体を90º回転させ重力方向を変化させると、根が重力方向に屈曲しにくいことが分かりました(図1)。
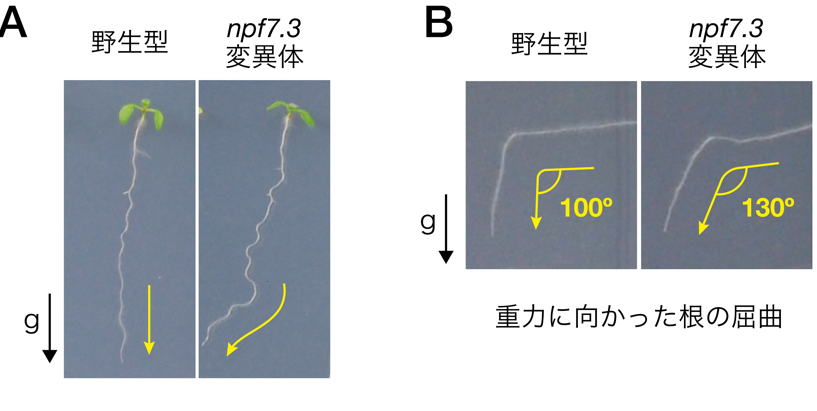 図1 NPF7.3の変異によるシロイヌナズナ根の重力屈性の異常
図1 NPF7.3の変異によるシロイヌナズナ根の重力屈性の異常
(A) 発芽後1週間栽培した野生型シロイヌナズナとnpf7.3変異体。野生型の根は重力方向に真っ直ぐに伸びたが、npf7.3変異体の根は左右に向かって不規則に伸びた。
(B) 野生型シロイヌナズナとnpf7.3変異体を90°回転させ、根にかかる重力方向を変えてから、1日後に根の屈曲を観察した。野生型の根はほぼ直角(90〜100°)に屈曲し重力方向に伸びたが、npf7.3変異体の根は重力方向に屈曲しにくかった。
*黒矢印は重力方向を指す。
植物の重力応答にはIAAが重要な役割を果たしていることから、NPF7.3がIAAもしくはその前駆体の細胞内取り込み輸送体であると予想されました。そこで、酵母細胞を用いて、IAAおよびIBAに対する輸送活性を調べたところ、NPF7.3はIAAよりもIBAを効率良く細胞内に取り込むことが分かりました(図2)。また、LC-MS[9]を用いた分析により、npf7.3変異体の根に含まれるIBA量は野生型の半分程度であることが明らかになりました。
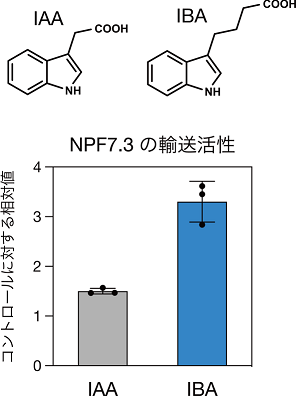 図2 酵母細胞を用いたNPF7.3のIBA取り込み活性
図2 酵母細胞を用いたNPF7.3のIBA取り込み活性
上:インドール酢酸(IAA)とインドール酪酸(IBA)の構造。
下:NPF7.3を発現した酵母細胞は、IAAよりもIBAを積極的に細胞内に取り込むことが分かった。
次に、npf7.3変異体における重力変化に応答したオーキシン(IAA)不等分布の形成を野生型と比較しました。その結果、オーキシン応答性マーカーであるDR5rev:GFP遺伝子[10]を導入したnpf7.3変異体では、野生型で見られる重力側でのGFP蛍光の偏りが著しく阻害されることが分かりました(図3)。これらの結果から、NPF7.3はIBAを細胞内へと取り込み、取り込まれたIBAがIAAへ変換されることで、根端の重力応答が誘導されていると考えられます。
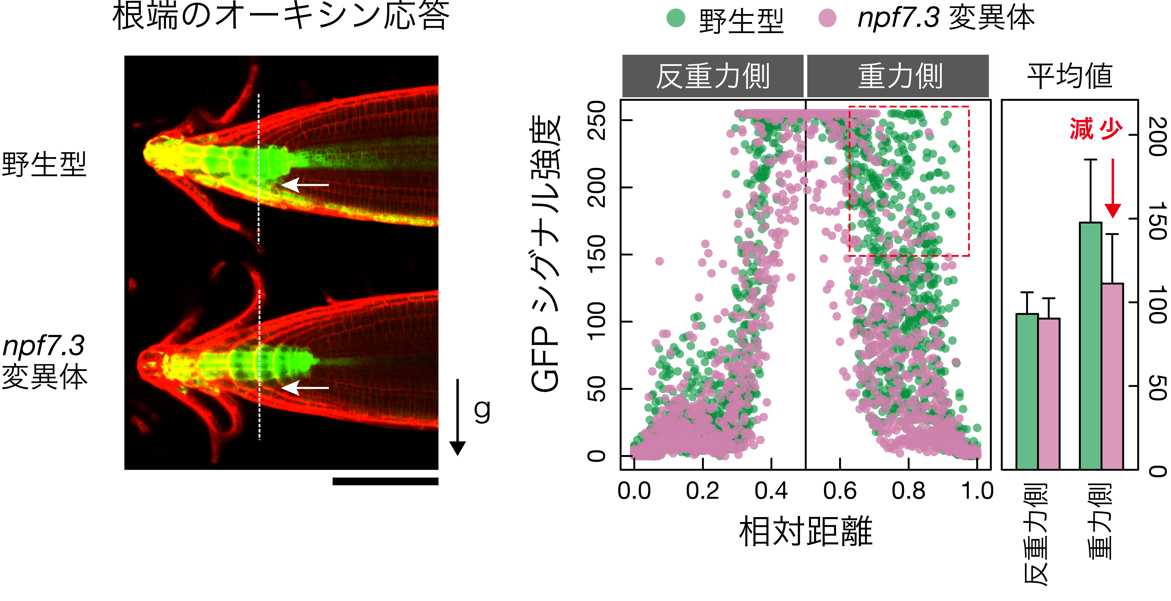 図3 重力刺激に応答した根端のオーキシン(IAA)不等分布形成
図3 重力刺激に応答した根端のオーキシン(IAA)不等分布形成
左: オーキシン応答性マーカー遺伝子(DR5rev:GFP)を導入した野生型とnpf7.3変異体を90°回転させ、根にかかる重力方向を変化させてから4時間後にGFP蛍光を観察した。白矢印で示すように、野生型では重力側に偏ってオーキシン応答(GFP蛍光)が強く誘導されたが、npf7.3変異体ではその応答が著しく阻害された。黒矢印は重力方向を指す。スケールバーは100マイクロメートル(μm、1μmは1000分の1mm)。
右: 左画像の白点線上のGFPシグナル強度と相対距離を示したグラフ(左)とGFPシグナル強度平均値のグラフ(右)。npf7.3変異体では、重力側のGFPシグナル強度が野生型の約70%まで減少した。
3.今後の期待
本研究は、IBAの細胞内取り込み輸送体を新たに同定しただけでなく、これまで理解が進んでいなかった重力屈性におけるIBAの重要性を明らかにしました。IBAは、IAAとは異なる経路で輸送されていることが予想されています。今後、IBAの輸送経路の全体像を明らかにし、IAAとIBAの流れを上手く利用することで、根の形態を人為的に制御できるようになると考えられます。このような技術は、少ない肥料や水で収量を増大させるといった環境負荷を低減した農業の実現に貢献します。
今回の研究成果は、国際連合が2016年に定めた17項目の「持続可能な開発目標(SDGs)[11]」のうち「2.飢餓をゼロに」と「15.陸の豊かさも守ろう」に大きく貢献すると期待できます。
4.論文情報
【タイトル】
The Arabidopsis NRT1/PTR FAMILY Protein NPF7.3/NRT1.5 is an Indole-3-butyric Acid Transporter Involved in Root Gravitropism
【著者名】
Shunsuke Watanabe, Naoki Takahashi, Yuri Kanno, Hiromi Suzuki, Yuki Aoi, Noriko Takeda-Kamiya, Kiminori Toyooka, Hiroyuki Kasahara, Ken-Ichiro Hayashi, Masaaki
Umeda, Mitsunori Seo
【雑誌】
Proceedings of the National Academy of Sciences of the United States of America (PNAS )
【DOI】
10.1073/pnas.2013305117
5.補足説明
[1] 屈性
生物が外部の刺激に応答して、一定の方向へ向かって成長あるいは旋回する性質のこと。刺激に向かって運動する正の屈性と刺激から遠ざかる負の屈性がある。
[2] オーキシン
最初に発見された植物ホルモン。葉、花、根など植物のさまざまな組織成長やパターン形成に重要な役割を持ち、光屈性や重力屈性を誘導するシグナル分子である。主要な天然オーキシンはインドール酢酸(IAA)で、根や茎の先端の分裂組織ではIAAが一定の方向に流れる(輸送される)ことで局所的に蓄積し、偏った成長が生じる。IAAは、主にアミノ酸であるトリプトファンからインドールピルビン酸を経て生合成される。これに対してイン ドール酪酸(IBA)は、微量なIAA前駆体でペルオキシソームβ酸化によりIAAに変換される。
[3] 輸送体
生体膜に局在するタンパク質であり、膜を貫通し孔を形成することで化合物の移動を仲介する。生体内の化合物の多くは、脂質二重膜である細胞膜や細胞内小器官の膜を通過できない。そのため、細胞間あるいは細胞内小器官と細胞質との物質交換には、それぞれに特別な輸送機構が必要と考えられており、輸送体はその一端を担っている。
[4] 植物ホルモン
植物が産生する生理活性・情報伝達を調節する機能を持つ物質のこと。植物に普遍的に存在し、低濃度で作用する、活性本体の化学構造や生理作用が明らかにされている物質が含まれる。オーキシン、ジベレリン、サイトカイニン、アブシジン酸、ジャスモン酸、サリチル酸、エチレン、ブラシノステロイド、ストリゴラクトンが広く知られている。最近では、フロリゲンやペプチドホルモンも植物ホルモンとして認識されている。
[5] 根端
植物の根の先端部分の総称。最先端部から上部に向かって根冠、根端分裂組織、未分化組織の順で構成される。根の重力屈性の要となる組織で、オーキシンが高濃度に存在しており、重力側の細胞にその蓄積が偏ることで、細胞伸長が抑制され根が曲がる。
[6] シロイヌナズナ
アブラナ科シロイヌナズナ属の一年草で、世界で最もよく利用されているモデル植物。ゲノムサイズが1.3億塩基対(ヒトの25分の1)と小さく、2カ月程度で世代交代するため遺伝学的な解析に適している。
[7] Nitrate transporter 1/ Peptide transporter Family (NPF)
硝酸・小ペプチド輸送体ファミリー。文字通り、硝酸や小ペプチドの膜通過を仲介しているタンパク質ファミリー。最近では植物ホルモンなど重要な化合物を輸送するNPFが多数同定されており、多機能的な輸送体ファミリーとして注目を集めている。植物に広く保存されており、シロイヌナズナには53種類のNPFが存在する。
[8] 側根
主根から枝分かれして伸びる根。二次根とも呼ばれる。種子から地中に向かって真っ直ぐ伸びる主根の内鞘細胞が、細胞増殖することで形成される。この形成誘導にもオーキシンが重要な役割を果たしている。
[9] LC-MS
高速液体クロマトグラフィー(LC)と質量分析計(MS)を組み合わせた化合物分析装置。LC部では化学的特性の違いを、MS部では質量の違いをもとに、目的の化合物を分離できる。そのため、さまざまな種類の化合物に対して、定性的かつ定量的な分析が可能である。
[10] DR5rev:GFP遺伝子
オーキシン応答性の遺伝子発現調節領域(DR5rev)とオワンクラゲ緑色蛍光タンパク質(GFP)遺伝子を融合したキメラ配列。この配列を持つ植物では、オーキシンに強く応答している組織や細胞でGFPが緑色蛍光を発するため、オーキシン分布の観察に広く用いられる。
[11] 持続可能な開発目標(SDGs)
2015年9月の国連サミットで採択された、持続可能でよりよい世界を目指す国際目標。17のゴールおよび169のターゲットで構成されており、国連加盟193か国が、2016年から2030年の15年間でこれらを達成することを目標としている。
6.発表者・機関窓口
<発表者> ※研究内容については発表者にお問い合わせください。
理化学研究所 環境資源科学研究センター
適応制御研究ユニット
ユニットリーダー 瀬尾 光範 (せお みつのり)
基礎科学特別研究員 渡邊 俊介 (わたなべ しゅんすけ)
TEL:045-503-9666(瀬尾) FAX:045-503-9665 E-mail:mitsunori.seo[at]riken.jp(瀬尾)
<機関窓口> *今般の新型コロナウイルス感染症対策として、理化学研究所では在宅勤務を実施しておりますので、メールにてお問い合わせ願います。
理化学研究所 広報室 報道担当
E-mail:ex-press[at]riken.jp
奈良先端科学技術大学院大学 企画・教育部 企画総務課 渉外企画係
TEL:0743-72-5026 E-mail:s-kikaku[at]ad.naist.jp
東京農工大学 企画課広報係
TEL:042-367-5930 E-mail:koho2[at]cc.tuat.ac.jp
岡山理科大学 入試広報部
TEL:086-256-8412 E-mail:kouhou[at]ous.ac.jp
※上記の[at]は@に置き換えてください。