
【概要】
奈良先端科学技術大学院大学(学長:小笠原直毅、奈良県生駒市)のバイオサイエンス研究科植物発生シグナル研究室 中島敬二(なかじま けいじ)教授らの研究グループは、京都大学、大阪市立大学、広島大学、神戸大学、近畿大学との共同研究により、植物が次世代をつくる際に必要な生殖細胞の形成を制御する遺伝子を発見しました。生殖細胞はコケなどでは卵や精子、種子植物では卵や花粉に当たりますが、この遺伝子は、すべての植物に存在しており、植物種を問わずに生殖細胞の形成に働いていると考えられます。植物において生殖細胞の形成を中心的に制御する遺伝子の発見は初めてのことです。今後は、この遺伝子を手掛かりに、植物が生殖細胞を作るメカニズムや、生殖細胞が受精後にさまざまな細胞に分化する多能性を獲得する仕組みを明らかにできると期待されます。
この研究成果は、アメリカ東部時間の6月23日(木)正午【プレス解禁日時:日本時間平成28年6月24日(金)午前1時】付けで、カレントバイオロジー(5-year impact factor, 10.1)のオンライン版に発表されます。
この遺伝子はRKDファミリーと呼ばれる一群の遺伝子で、遺伝子DNAの必要な遺伝情報の部分をメッセンジャーRNAという分子にコピー(転写)する際に、その働きを調節するタンパク質(転写因子)の遺伝子です。
中島教授らは、5年前にモデル植物であるシロイヌナズナにおいて、この遺伝子ファミリーのメンバーの1つであるRKD4と呼ばれる遺伝子が、受精後の胚の形成を制御することを明らかにしています。一方でシロイヌナズナのゲノムには、RKD1-RKD5の5つのRKD遺伝子が存在していますが、RKD4以外の4つの遺伝子の機能は未解明のままでした。また、RKD遺伝子は様々な植物のゲノムに広く存在していますが、これらの働きも分かっていませんでした。
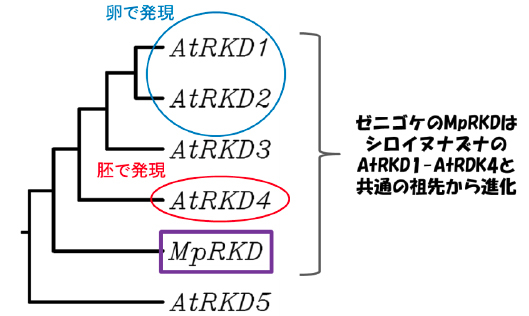
こうしたことから、中島教授らは、苔類(コケ植物)のゼニゴケ(Marchantia polymorpha)に注目しました。ゼニゴケはゲノム構成が単純で、かつ狙った遺伝子を容易に破壊できるという実験上の利点があります。さらに、ゼニゴケは陸生植物の進化の原点に位置しているうえ、1種のRKD遺伝子しか持たないことから、植物に共通したRKD遺伝子の役割を解明できるのではないかと考えて研究を進めました。
ゼニゴケはオスとメスに別れた雌雄異株の植物です。ゼニゴケのRKD遺伝子(MpRKD遺伝子)は、メス個体においては卵母細胞(卵子のもとになる細胞)と卵細胞、オス個体では精母細胞(精子のもとになる細胞)でそれぞれ強く発現していました。この遺伝子を破壊すると、植物の形態そのものには異常が見られないものの、生殖細胞(卵子と精子)の形成には重篤な異常が見られました。特に卵の異常は致命的で、細胞壁の消失や核の肥大化といった通常の卵細胞で見られる現象が起こらず生殖能が失われていました。
これらの結果は、ゼニゴケがもつ唯一のRKD遺伝子が、生殖細胞の形成に必須であることを示しています。種子植物のシロイヌナズナやコムギにおいても、複数の種類のRKD遺伝子が卵細胞で発現することが分かっており、RKD遺伝子による生殖細胞形成の制御は、植物の進化を通じて保存されたものであると考えられます。
この研究成果は、生殖細胞の形成を中心的に制御する因子を植物において初めて明らかにしたものです。今後はRKD遺伝子の作用機構を研究することで、生殖細胞の形成メカニズムや、分化多能性の獲得機構が明らかになることが期待されます。また、植物における生殖細胞形成の仕組みを明らかにすることで、作物の効率的な繁殖や育種技術の向上にも寄与することができると期待されます。
【論文情報】
タイトル:An evolutionarily conserved plant RKD factor controls germ cell differentiation
著者:Satoshi Koi, Tetsuya Hisanaga, Katsutoshi Sato, Masaki Shimamura, Katsuyuki T. Yamato, Kimitsune Ishizaki, Takayuki Kohchi and Keiji Nakajima
掲載誌:Current Biology (Cell Press)
掲載日時(オンライン版):2016年6月23日(木)正午(アメリカ東部時間)
naistar:http://hdl.handle.net/10061/10652
(NAIST Academic Repository:naistar)
【解説】
有性生殖は個体の遺伝情報を混ぜ合わせて子孫の性質に多様性をもたらし、種の維持を図るプロセスであり、生物進化の原動力です。有性生殖の成功には、卵と精子という非常に特殊化した細胞を作る過程が必須です。生殖細胞は、細胞同士の融合(受精)と、それに続く核内での染色体の統合、という2つの過程を経て接合子となり、胚を形成します。動物の生殖細胞は、個体発生のごく初期に取り分けられた細胞から作られます。これに対し、植物の生殖細胞は個体発生の最後に、花などの生殖器官内において、分化した細胞が初期化(リプログラミング)されることにより作られます。ところが、植物において、分化した体細胞から卵や精子という受精能や分化多能性を持つ細胞が作られるメカニズムは未解明のままです。
モデル植物のシロイヌナズナにおいて、RKD4遺伝子が体細胞の初期化と胚発生を制御していることが知られています(2011年7月にプレスリリース)。RKDは植物に広く存在する遺伝子ファミリーで、転写因子をコードしています。シロイヌナズナのゲノムにはRKD4以外にも4つのRKD遺伝子があり、それぞれが様々な植物種の生殖器官で発現していることが分かっており、植物の生殖に共通した重要な機能を担っていると考えられていました。しかし、それを裏付ける証拠は示されていませんでした。
ゼニゴケは、雌雄異株の苔類(コケ植物)の一種であり、陸上に生育する植物の祖先型です。京都大学の河内孝之(こうち たかゆき)教授らにより、全ゲノムの解読をはじめ、形質転換や遺伝子破壊などの研究技術が整備され、新たなモデル植物として急速に普及しつつあります。ゼニゴケの形態は種子植物とは大きく異なりますが、生殖細胞の形成機構には共通点があると考えられます。
また、ゼニゴケは有性生殖だけでなく、無性芽(むせいが)を介して無性生殖でも増殖します。このため、生殖細胞が作られなくなった変異体でも、系統を維持して研究材料にすることができます。さらにゼニゴケはゲノム中に機能が重複した複数の遺伝子をほとんど持っていません。実際、シロイヌナズナが5つのRKD遺伝子を持つのに対して、ゼニゴケは1つのRKD遺伝子(MpRKDと命名)しか持っておらず、MpRKD遺伝子は、陸上植物の進化を通じて保存されたRKD遺伝子の祖先的な機能を保持していると考えられます(図1)。

【実験の結果】
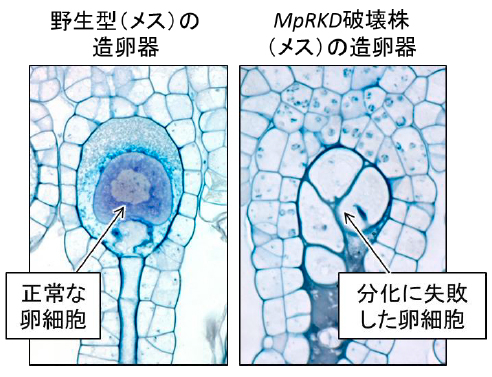
ゼニゴケの生活環を通じてMpRKD遺伝子の発現を調べたところ、MpRKDはメス個体では卵母細胞および成熟した卵細胞で、オス個体では精母細胞で強く発現していました。ゼニゴケの卵細胞は、破れた傘のような形をした雌器托(しきたく)の下にぶら下がった造卵器(ぞうらんき)の内部に分化します(図2、図3左)。卵細胞は他の細胞と比較して圧倒的に大きく、受精の障害となる細胞壁が消化されるために球形をしています(図3左)。また、細胞質に富み核が巨大化しています(図3左)。
MpRKDを破壊した実験では、このような卵に特徴的な細胞分化が起こらず、異常な細胞分裂を生じ、細胞の液胞化が起こっていました(図3右)。さらに破壊株の造卵器は精子を誘引できませんでした。
一方、MpRKDを破壊したゼニゴケのオス個体では、精子形成のもとになる精母細胞の形成に異常が見られました。野生型のゼニゴケのオス個体では、平らな傘状の雄器托(ゆうきたく)の内部にある造精器(ぞうせいき)の中で精母細胞が同調的に増殖し、特異的な変態過程を経て運動性のある大量の精子が作られます。遺伝子を破壊した株の造精器内では、精母細胞の分裂が同調せず、崩壊している細胞が多く見られました。しかしメスの卵とは異なり、一部の精子は正常に形成されていました。
以上の結果から、ゼニゴケにおいてMpRKD遺伝子が、オスとメスに共通して生殖細胞の分化を担う制御因子であることが確かめられました。シロイヌナズナにおいても、RKD1やRKD2遺伝子が卵細胞で強く発現していることが分かっていましたが、これらが卵細胞の形成を制御していることを示すことは困難でした。こうした困難さは遺伝子の数の問題だけではなく、種子植物では卵細胞が小さく、かつ蕾に包まれた雌しべの内側にある胚珠の中で分化するため、その細胞の形態や細胞内部を明瞭に観察することが不可能であることによります。
RKD遺伝子の機能としては、シロイヌナズナでRKD4が胚発生を制御することが先に発見されましたが、この機能は、むしろ進化の過程でRKD遺伝子の数が増えてから獲得されたものであり、生殖細胞の形成を制御することが、祖先的な機能であると考えられます。
この研究成果の特徴は、植物において生殖細胞の形成を中心的に制御する遺伝子を初めて発見したことです。それに加え、既存のモデル植物では不可能であった研究を、ゼニゴケという新しいモデル植物の特徴をうまく利用して明らかにしたことも特筆すべき点です。
本研究は、平成23年度 JSTさきがけ採択課題「植物生産能の高度利用に向けた「植物iPS遺伝子」の応用展開」、科学研究費補助金、新学術領域研究「植物発生ロジック」および基盤研究(B)、倉田奨励金の支援により行われました。
【今後の発展】
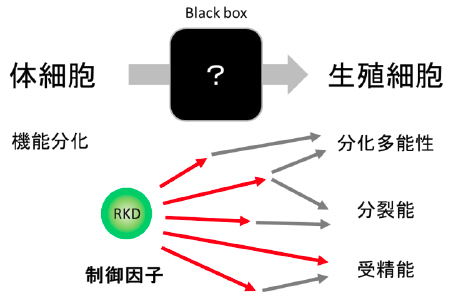
これまで植物が生殖細胞を作り出す仕組みや、それにかかわる遺伝子はほとんど見つかっていませんでした。これは生殖細胞が非常に小さく、植物体の奥深くで作られるために解析が難しかったためです。ゼニゴケは、大きな生殖細胞を比較的体外に近い場所で作るため、観察するのが容易です。また今回、生殖細胞の形成を司るマスター制御因子を捕まえることが出来ましたので、これを手掛かりに生殖細胞を作る実働遺伝子を芋づる式に捕まえることが出来るようになります(図4)。
また、RKD遺伝子を使うことで、分化した体細胞から人工的に生殖細胞を作ることが出来るかもしれません。これまでRKD4や他の転写因子を使って、体細胞から胚を作ることは出来ていました。こうしてできた胚(不定胚と呼びます)は元の細胞と同じ遺伝情報を持つため、優良な株のクローンを大量に生産するのに適しています。しかし作物の育種においては、異なる親個体をかけ合わせて多様な子孫を作り出し、それらの中から優良な個体を選抜する方法がとられます。発芽から生殖までの時間が長ければ、育種に膨大な時間がかかります。芽生えの細胞から生殖細胞を作ることができれば、育種にかかる時間を大幅に短縮できます。もちろん生殖細胞を試験管内で受精させる技術を同時に開発する必要があります。
【用語解説】
・RKD遺伝子(MpRKD遺伝子)
RKD遺伝子は、様々な植物種のゲノムに広く存在する遺伝子で、動物には存在しません。RKD遺伝子は、転写因子(ゲノムDNAからメッセンジャーRNAへの転写を組織や発生段階特異的に制御するタンパク質)をコードしています。植物には多くの転写因子ファミリーがありますが、RKDファミリーは、比較的最近になって発見され、陸上植物が必ず持っているにも関わらず、その働きはほとんど知られていません。種子植物のゲノムには複数のRKD遺伝子が存在していますが、これらは共通した機能を担っていると考えられ、破壊株を用いて働きを調べるのは困難です。これに対し、ゼニゴケのゲノムにはRKD遺伝子は1つしかなく、ゼニゴケの学名(Marchantia polymorpha)の頭文字をとって、MpRKD遺伝子と命名しました(図1)。
・シロイヌナズナの生殖
種子植物の生殖細胞はやや複雑な過程を経て形成されます。まず、めしべの中にある胚珠(将来種子になる微小な袋)の1細胞が減数分裂し、染色体数が半減した細胞が4個作られます。これらのうち3つは退化し、残りの1つが(染色体数が半数のまま)3回分裂して、8個の細胞を作ります。これらのうち1つが卵細胞になります。また残りの7個のうち2個が融合して中心細胞が作られます。一方、おしべの先端にある葯では多数の細胞が減数分裂し、染色体数が半減した細胞が4個作られます。これらがそれぞれ花粉へと分化します。花粉の中では核が分裂し、2つの精細胞と1つの花粉管核が作られます。花粉がめしべの先端の柱頭に付着すると、花粉の一部が突出してチューブ状の花粉管を伸ばします。これがめしべ内の胚珠まで達すると、先端から2つの精細胞が放出されます。これらのうち1つは卵と受精して胚を作ります。胚は次世代の個体となります。もう1つの精細胞は中心細胞と受精して胚乳を作ります。胚乳は胚やめばえの栄養貯蔵を行います。白米はイネの胚乳であり、胚は精米時に除去されます。
・ゼニゴケの生殖(図2)
ゼニゴケの生活環は、種子植物とは大きく異なっています。まず減数分裂で生じた半数体細胞(相同染色体を1本ずつしか持たない細胞)が個体を作っています(進化的にはこちらが先です)。また雌雄異株であり、X染色体をもつ株がメス、Y染色体をもつ株がオスになります。ゼニゴケは地面を覆うように葉状体を作り、条件が整うと生殖器官を形成します。メスの生殖器官は破れ傘のような形をしており、その下側に逆さのフラスコ型の造卵器を多数つけます。造卵器の中に卵細胞が1つずつ形成されます。オスの生殖器は平らな傘のような形をしており、その内部に多数の造精器をつけます。1つの造精器の中で数万の精子が同調的に形成されます。精子はベン毛を使って泳ぐことが出来るため、降雨などで造精器が濡れると外部に漏出し、未知のシグナルに誘引されて造卵器へと移動します。卵に達した精子は受精し、相同染色体を2本ずつもつ接合子となります。接合子が分裂して胚を形成しますが、種子植物とは異なり、胚は成体を形成することなく減数分裂し、半数体の胞子を形成します。胞子にはXまたはY染色体が分配されるので、オスとメスの胞子が同数出来ます。胞子はやがて発芽して葉状体を形成します。
ゼニゴケは無性生殖によっても増殖できます。この場合は、オス・メスともに葉状体の中央にカップ状の器官(杯状体)ができ、その中に多数の無性芽が形成されます。これらが地面に散乱すると、それぞれが分裂して新たな葉状体となります。葉状体は親植物と同じ遺伝情報を持つクローンです。ゼニゴケは無性生殖でも繁殖できるため、生殖細胞に異常を持つ破壊株を維持して解析に供することが出来ます。