
【概要】
奈良先端科学技術大学院大学(学長:小笠原直毅、奈良県生駒市)のバイオサイエンス研究科植物発生シグナル研究室 中島敬二(なかじま けいじ)教授らの研究グループは、理化学研究所との共同研究により、植物の根の先端を覆うキャップ型の根冠(こんかん)組織から、生きた細胞が自発的に剥がれ落ちる仕組みを明らかにしました。根冠は根が土壌中を成長する際に、根の先端を保護したり、重力方向を感じたり、土壌の環境を調整したりする重要な組織です。根冠細胞の剥離は、植物の成長のみならず、植物による土壌への働きかけにも大きな影響があり、その仕組みの解明は、植物の成長力の改良や土壌環境の改善にも資する研究成果です。
この研究成果は、英国時間の11月1日(火)【プレス解禁日時:日本時間平成28年11月1日(火)午前9時】付で、ディベロップメント(英国発行の発生生物学の専門誌)のオンライン版に発表されます。
中島教授らの研究グループは、モデル植物であるシロイヌナズナを実験材料に用い、根冠(※1)の分化を調節する転写因子(遺伝子からmRNAへの転写を活性化したり抑制したりするタンパク質)(※2)として知られていた、SMB、BRN1、BRN2の働きを詳細に調べました。これらの転写因子は、根冠内で働き、根冠の分化を調節することが報告されていましたが、根冠のどこで、いつ、どのような遺伝子の調節を行っているかは分かっていませんでした。
実験の結果、これらの調節因子は、過去の報告に反して根冠の中で空間的に使い分けされていることが分かりました。SMBが根冠全体で働いているのに対し、BRN1とBRN2は、「細胞が根の最も外側の位置にある」ことを感知して働き、これによって根冠の外側の細胞がもつ特殊な機能を調節することが分かりました。
次にこれらの転写因子が調節する標的遺伝子を探索し、根冠の形成や働きに関与していると思われる60個の遺伝子を見つけました。これらの中には、植物の細胞壁に作用する酵素や、脂質を合成する酵素、細胞外へ物質を分泌するさいに働くタンパク質などが含まれており、これまでは現象の記述にとどまっていた根冠の機能を、遺伝子・分子のレベルで解析出来るようになりました。
中島教授らは、まず細胞壁の主成分のペクチンを分解する酵素であるポリガラクツロナーゼ(Polygalacturonase = PG)の遺伝子に着目し、これをRCPG (Root cap polygalacturonase) と名付けて詳細に解析しました。細胞壁の主成分の1つであるペクチンは、隣接する細胞をつなぎ留める働きを持つことに着目したのです。実験の結果、RCPGタンパク質は、根の最外層に位置する根冠細胞で急激に作られ、自身の細胞壁を分解して根から切り離していることが分かりました。これにより、根冠の外側の細胞が、生きたまま、自発的に剥がれてゆく仕組みが遺伝子・タンパク質のレベルで明らかになりました。
根冠細胞は根の成長を支えるのみならず、剥離した細胞が土壌中に散布されることで、土壌環境を整えたり、大気中の炭素を土壌へと還流する役割を担っています。根冠細胞が、生きたまま自発的に根から外れてゆく、という一見奇異な現象の裏には、このような生態学的に重要な意義が隠されていると考えられます。今回の成果は、植物の根冠細胞がもつユニークな機能が発現する仕組みを、遺伝子・タンパク質レベルで明らかにしたものですが、将来は植物の成長制御や土壌環境の保全等にもつながる研究成果です。
【論文情報】
タイトル:Control of root cap maturation and cell detachment by BEARSKIN transcription factors in Arabidopsis
著者:Masako Kamiya, Shin-ya Higashio, Atsushi Isomoto, Jong-Myong Kim, Motoaki Seki, Shunsuke Miyashima, and Keiji Nakajima
掲載誌:Development (The Company of Biologists, 英国に拠点をもつ生物学論文誌の出版社)
http://dev.biologists.org/content/143/21/4063.full
掲載日時:2016年11月1日(火)オンライン版
naistar:http://hdl.handle.net/10061/11095(NAIST Academic Repository:naistar)
【解説】
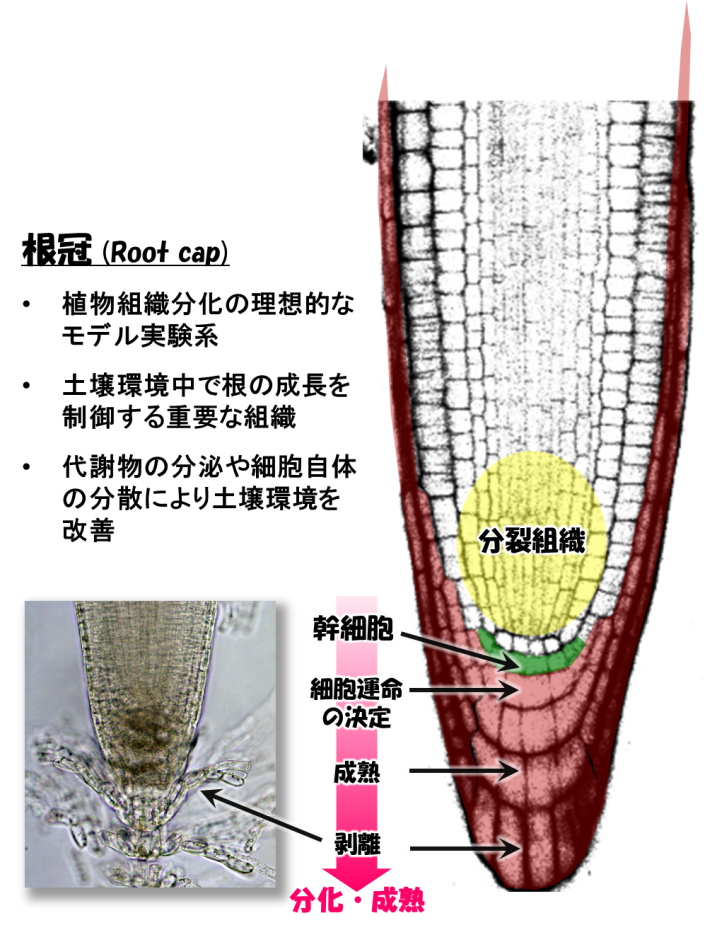
植物の根の先端には根冠(こんかん、Root cap、※1)と呼ばれる組織がはまっています。根冠は、根端の分裂組織を土壌の刺激から物理的に保護するのみならず、重力の方向を感知して根の伸びる方向を調節したり、土壌中に様々な物質を分泌して微生物の生育をコントロールしたり、など、植物の成長と土壌環境の改善に必須の役割を担っています(図1)。
根冠組織は、植物種により数層から数十層の細胞層からできていますが、その作られ方には植物の他の組織には見られないユニークな特徴があります。それは、根冠組織の最も内側で新しい細胞層が生み出され、最も外側の細胞層から順に剥落してゆくことです(図1)。このように、根冠を形作る細胞群は常に新しい細胞へと入れ替わっています。これは、あたかもヒトの皮膚の細胞と似ていますが、根冠が皮膚と異なるのは、外側の細胞が一定の時間間隔で、自発的に、しかも生きたまま剥落することです。根冠組織の中で外側にある細胞だけが、どのようにして剥離してゆくのか、その仕組みはこれまで分かっていませんでした。
【実験の結果】
中島教授らの研究グループは、モデル植物であるシロイヌナズナを実験材料に用い、まず3つの転写因子(遺伝子からmRNAへの転写を活性化したり抑制したりする調節タンパク質、※2)であるSMB(ソンブレロ)、BRN1(ベアスキン1)、BRN2 (ベアスキン2)を手掛かりに研究を始めました。これらの転写因子は、オランダの研究グループや、中島教授らのグループにより、根冠の分化を制御する鍵因子として見つかっていたものです(※3)。しかし、これまでこの3つの転写因子がどのように使い分けられ、どのような遺伝子の転写を調節しているのかは、分かっていませんでした。
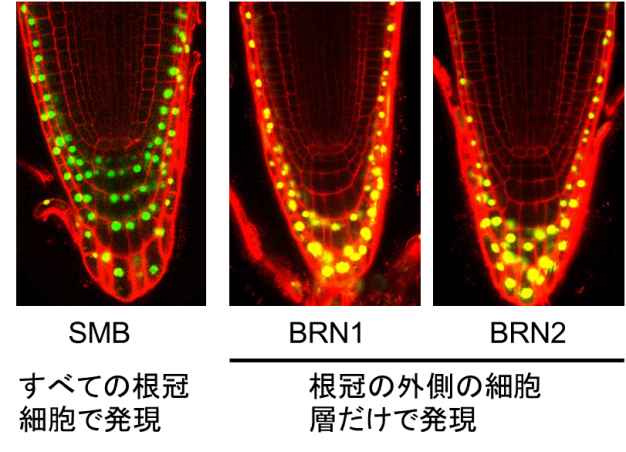
まず、この3つの転写因子が根冠のどこで働いているかを調べたところ、SMBが根冠全体で働いているのに対し、BRN1とBRN2の2つは、根冠の最も外側にある1-2層の細胞層のみで働いていることが分かりました(図2)。またBRN1とBRN2は、「細胞が根の最も外側にある」ことを感知して働くことが分かりました。さらにSMB, BRN1, BRN2が、遺伝子からmRNAへの転写を活性化する因子であることを、タバコの培養細胞を用いた実験で明らかにしました。
次にこれら3つの転写因子が調節している標的遺伝子を探索しました。これには、①野生型植物、②3つの転写因子が働かない変異体、③3つの転写因子が野生型よりも強く働く植物体、の3種類の植物で、全遺伝子の発現量を比較し、②<①<③の順で発現が多い遺伝子を探す方法を用いました。その結果、これら3つの転写因子によって調節され、根冠の形成や働きに関与していると思われる遺伝子を60個見つけました。これらの中には、植物の細胞壁に作用する酵素や、脂質を合成する酵素、細胞外への分泌に働くタンパク質などが多く含まれていました。
中島教授らは、細胞壁に作用する酵素を作る遺伝子の中から、細胞壁の主成分の1つであるペクチン(※4)という糖のポリマーを分解する酵素であるポリガラクツロナーゼ(Polygalacturonase = PG)を作る遺伝子に着目し、これをRCPG (Root cap polygalacturonase) と名付けました。ペクチンは細胞壁においてセルロース繊維の間を充てんするゲル状の物質で、隣り合った細胞どうしを接着する働きをもっています。ポリガラクツロナーゼ(※5)はペクチンの主鎖であるポリガラクツロン酸のポリマーを加水分解して切断する酵素です。
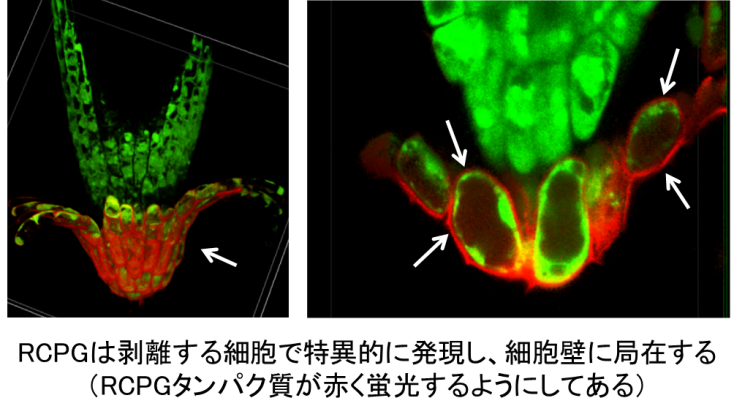
まずRCPGが根冠のどこで、いつ働くのかを調べたところ、RCPGタンパク質は、最外層に位置する根冠細胞で急激に作られて、それらの細胞壁へと運ばれることが分かりました(図3)。一方で、より内側の根冠細胞や、根の他の組織では、ほとんど働かないことが分かりました。この働き方は、BRN1とBRN2転写因子によく似ていたため、それらとの関係を調べたところ、BRN1とBRN2が働かなくなっている変異体ではRCPGも働かなくなっていました。またクロマチン免疫沈降(※6)という方法により、少なくともBRN1がRCPG遺伝子の調節領域に直接結合し、その転写を活性化していることが分かりました。
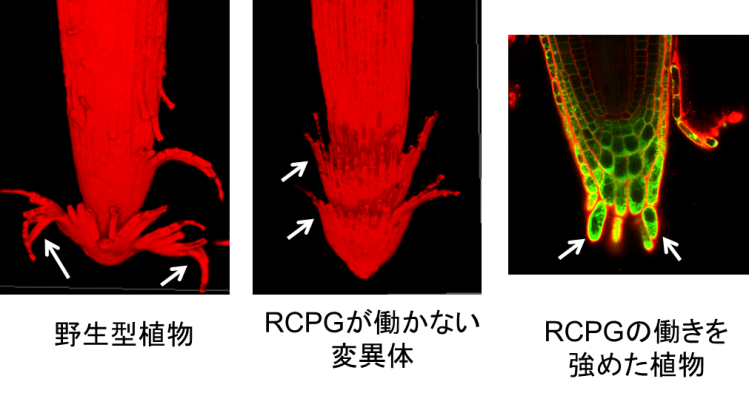
RCPG遺伝子を破壊した変異体植物を作成し、根冠の剥がれ方を野生型と比較したところ、野生型では根冠の最外層が、大きく反り返って剥がれ落ちていたのに対し、変異体においては、根冠の最外層がカップ状の外形を維持して、根の先端にはまったままになっていました(図4)。反対にRCPGの働きを強めた植物を作成したところ、その根冠の最外層は層状に剥がれず、1つ1つの細胞が根の先端からバラバラと剥がれ落ちていました(図4)。
以上の結果から、①根冠の外側にある細胞が根の表面にいることを感知し、②その情報をもとにBRN1とBRN2の2つの転写因子を活性化させ、③これらがRCPG遺伝子を活性化してペクチン分解酵素を作り、④生成した酵素が最外層の細胞の細胞壁に運ばれてペクチンを分解し、⑤細胞どうしの接着が緩んで生きたまま細胞が剥離し、⑥外から2層目にあった細胞が最外層となって①のステップに戻る、という一連の流れを明らかにすることができました。また、RCPGの働きを特定の細胞だけに限定し、さらにその量を適度に調節することで、一番外側の細胞が層としての形を保ったまま、順に剥がれてゆく仕組みを明らかにすることができました。
こうした仕組みによって根冠細胞が生きたまま剥がれることで、細胞内にたまった様々な代謝産物が分解されることなく、直接土壌中に散布されます。それによって土壌の生物学的あるいは化学的環境が整えられます。根冠が根の先端で整えた環境中を根の残りの部分が伸長してゆくことで、栄養分の効率的な吸収が促進され、また植物にとって有害な病原菌の繁殖が抑えられることが想像されます。
【今後の発展】
根冠細胞は根の成長を支えるのみならず、剥離した細胞が土壌中に散布されることで、土壌環境を整えたり、大気中の炭素を土壌へと還流する役割を担っています。植物の根の先端にこのような重要な組織が存在することは、一般にはあまり知られていませんが、最近では高校生物の教科書にも掲載されるようになっており、その特殊な機能が発現する仕組みを理解してゆくことは、今後非常に重要になってくると思われます。本論文では、根冠の機能のうちでも特に細胞の剥離が調節される仕組みに焦点を絞って実験を行いましたが、今回見つかった60個の遺伝子の中には、他にも興味深い遺伝子が多数残っており、今後はこれらの働きを順次解明してゆくことで、根冠という植物の成長に重要な組織がどのように働いているのかを明らかにできることが期待されます。また土壌中での根と微生物の相互作用には未知の部分が多いですが、根冠の機能を解明することで、生態学的な観点からも新しい知見が得られる可能性があります。
【用語解説】
※1 根冠
根の先端にペンのキャップのようにはまっている組織。植物の根には必ず存在します。根冠は根端にある分裂組織(根を伸長させるのに必要な活発に分裂する細胞の集まり)を覆って物理的に保護しています。根冠の中央には重力感受細胞があり、この細胞中で平衡石(デンプンを蓄えた比重の重い細胞内小器官)が沈むことで重力方向を感じています。根冠の一番外側の細胞は分泌機能に特化しており、土壌中に代謝産物を分泌していると考えられています。最外層細胞の一部はプログラム細胞死により自殺するが、多くは生きたまま剥離して土壌中に分散します。細胞内の有機物はやがて微生物に利用されるため、根冠の剥離は大気から土壌へ炭素を還流させる役割も担っています。
※2 転写因子
遺伝子DNAの調節領域に結合して、mRNAへの転写を調節するタンパク質です。転写の活性化因子と抑制因子があります。シロイヌナズナのゲノムには約2000個の転写因子がコードされています。転写因子ごとに、働く細胞やタイミング、標的とする遺伝子が異なっているため、これらの転写因子の働き方の組み合わせで、細胞ごとの性質や働きの違いが現れます。転写因子を作る遺伝子自体も転写因子により調節され、また複数の転写因子が複合体を作って働くため、その調節ネットワークは非常に複雑化しています。
※3 SMB, BRN1, BRN2転写因子
植物に特異的なNAC(ナック)転写因子ファミリーに属します。シロイナズナのゲノムにはNAC転写因子をコードする遺伝子が約100個存在し、植物の発生や生理機能の様々な局面を調節しています。SMB, BRN1, BRN2の3つは根冠で特異的に機能し、根冠細胞の分化を調節していることが分かっていましたが、この3つがどのように使い分けられ、どのような遺伝子を調節しているかは、これまで分かっていませんでした。
※4 ペクチン
植物の細胞壁を構成する主要な糖鎖ポリマーの1つです。様々な単糖がつながってできていますが、主鎖を構成するのは、ガラクツロン酸とラムノースと呼ばれる単糖です。ペクチンはポリマーどうしがカルシウムにより架橋されてゲル状の性質を示します。ペクチンは一次細胞壁のセルロース繊維の間を充てんすることで、隣り合った細胞どうしを接着する働きがあります。
※5 ポリガラクツロナーゼ
ペクチンを分解する酵素(ペクチナーゼと総称される)の1つで、ガラクツロン酸のポリマーを加水分解して断片化します。シロイヌナズナのゲノムにはポリガラクツロナーゼをコードする遺伝子が60個以上あり、それぞれが異なった組織やタイミングで働いています。
※6 クロマチン免疫沈降
ゲノムDNAに結合する転写因子などのタンパク質が、細胞核内でゲノムDNAのどこに、どれくらい結合しているかを調べる実験手法です。細胞核から抽出した転写因子などの目的タンパク質を、それに結合しているDNAと一緒に特異的な抗体で精製し、結合した特定のDNAをPCRで検出したり、あるいは結合したDNAすべてを次世代シーケンサーで分析したりすることで、結合サイトや結合量を知ることができます。
【本研究内容についてコメント出来る方】
東京大学大学院理学系研究科
塚谷 裕一(つかや ひろかず) 教授
Tel:03-5841-4047 Email:tsukaya@bs.s.u-tokyo.ac.jp
神戸大学大学院理学研究科
深城 英弘(ふかき ひでひろ)教授
Tel: 078-803-5721 Email: h-fukaki@port.kobe-u.ac.jp
【本プレスリリースに関するお問い合わせ先】
奈良先端科学技術大学院大学 バイオサイエンス研究科 植物発生シグナル研究室
中島 敬二(なかじま けいじ)教授
TEL:0743-72-5560(オフィス) FAX:0743-72-5569(オフィス)
E-mail:k-nakaji@bs.naist.jp