
ゼニゴケは遺伝子のオモテとウラを使ってメスとオスを作り分けている
~ 性差を生み出す巧妙な「裏ワザ」が明らかに~
【概要】
奈良先端科学技術大学院大学(学長:横矢直和、奈良県生駒市)先端科学技術研究科 植物発生シグナル研究室の中島 敬二(なかじま けいじ)教授と、京都大学(総長:山際壽一、京都市左京区)大学院生命科学研究科 遺伝子特性学研究室の河内 孝之(こうち たかゆき)教授の研究グループは、近畿大学、広島大学、豪Monash大学との共同研究により、陸上植物に共通した性分化制御遺伝子FGMYBを発見しました。またゼニゴケは、FGMYB遺伝子をつくるDNA二本鎖の表側と裏側を巧妙に使い分け、これを雌雄の性差を生み出すスイッチとして利用していることも明らかにしました。
研究グループは、ゼニゴケ(Marchantia polymorpha)とシロイヌナズナ(Arabidopsis thaliana)のメスの生殖器官の遺伝子発現を比較しました。ゼニゴケは陸上植物の進化の系統樹の中で最も基部に位置するコケ植物であり、シロイヌナズナは最先端部に位置する被子植物です。研究グループは両植物種のメスの生殖器官で共通してはたらく遺伝子を23組見つけ出し、それらの中からFGMYB遺伝子に着目しました。FGMYBは、他の遺伝子群のはたらきを統御するMYB型転写因子の1つをコードしています。シロイヌナズナのFGMYB (AtFGMYB)は、めしべの中で卵細胞を作り出す胚嚢の発達とはたらきを制御することが既に報告されていました。そこでゼニゴケのFGMYB (MpFGMYB)もメスの生殖機能に関係すると予想し、MpFGMYB遺伝子を破壊した変異体(Mpfgmyb変異体)を作成して解析しました。
ゼニゴケは保有する性染色体により雌雄の個体が分かれています。ところがMpFGMYB遺伝子を破壊した株(Mpfgmyb変異体)では、性染色体レベルではメスであるはずの個体がオスへと性転換していました。また、MpFGMYB遺伝子はメスとオスの両方が持っていましたが、メスだけではたらいていました。そこでオスにはMpFGMYBのはたらきを抑える機構があると考え、さらに解析を進めた結果、MpFGMYB遺伝子の裏側(対になったDNA二本鎖のうちMpFGMYBをコードしない側のDNA鎖)から、オスのみでmRNAが作られていることを発見し、これをSUFと名付けました。SUFを破壊した変異体を作成したところ、オスにおいてもMpFGMYBがはたらき、性染色体レベルではオスであるはずの個体がメスへと性転換していました。
以上の結果から、①FGMYBが陸上植物に共通したメス分化の促進遺伝子であること、また、②ゼニゴケにおいてはMpFGMYBと、その裏側のSUFから成る遺伝子領域が、雌雄を切り換えるスイッチとしてはたらいていること、が明らかとなりました。FGMYBは転写因子をコードしており、性差を作る遺伝子群のはたらきを調節していると考えられます。今後はFGMYBを手掛かりとし、植物が性に応じて特異的な形態や機能を発現する機構の解明が加速されることが期待されます。
本研究成果は陸上植物に共通した性分化の制御因子を世界で初めて同定したのみならず、それを用いた特異な性の切り換えスイッチの存在を明らかにしたものであり、有性生殖の成り立ちと進化を解明する上で学術的に大きなインパクトを持ちます。また植物に共通した性分化制御因子の発見は、作物の生殖能の改善や育種の効率化を目指した応用研究の展開にもつながる研究成果です。
本研究成果は、新学術領域研究「植物発生ロジックの多元的開拓」(領域代表者:塚谷裕一)、科研費基盤S「陸上植物の性分化:遺伝的頑健性と可塑性のメカニズム」(研究代表者:河内孝之、分担者:中島敬二、大和勝幸)ほか、以下の複数の研究グラントの支援により得られたものです。 MEXT-KAKENHI(25113007, 17H05841, 25113009); JSPS-KAKENHI (17J08430, 18K06285, 17H07424); AustralianResearch Council (DP170100049)
この研究成果は、現地時間の1月4日(金)正午付で、The EMBO Journal(エンボジャーナル、欧州分子生物学機構(EMBO)の学術誌、Impact Factor= 10.6)のオンライン版に発表されました。
【論文情報】
タイトル:
A cis-acting bidirectional transcription switch controls sexual dimorphism in the liverwort
(シスに働く二方向性の転写スイッチがゼニゴケにおける性の二形性を制御する)
著者:
Tetsuya Hisanaga1†, Keitaro Okahashi2†, Shohei Yamaoka2, Tomoaki Kajiwara2, Ryuichi Nishihama2, Masaki Shimamura3, Katsuyuki T. Yamato4, John L. Bowman5, Takayuki Kohchi2¶, Keiji Nakajima1¶
(久永哲也1†、岡橋啓太郎2†、山岡尚平2、梶原智明2、西浜竜一2、嶋村正樹3、大和勝幸4、ボーマン・ジョン5、河内孝之2¶、中島敬二1¶)
1奈良先端科学技術大学院大学、2京都大学、3広島大学、4近畿大学、5Monash大学
†共同第1著者、¶共同責任著者
掲載誌:
The EMBO Journal (欧州分子生物学機構(EMBO)の学術誌、EMBO Press (ドイツ)が、年24号を発行)
掲載日時:
2019年1月4日(金)正午 オンライン版
DOI:10.15252/embj.2018100240
naister:http://hdl.handle.net/10061/13097(NAIST Academic Repository:naistar)
【背景】
有性生殖はメスとオスの遺伝情報を混ぜ合わせ、多様な子孫を生み出すことで生物の進化を可能にしています。有性生殖で繁殖する生物は一般にメスとオスの形態が異なり(性の二形性)、それぞれの生殖器官から卵または精子という生殖細胞が作られます。性の二形性は生物進化の過程で独立に獲得されて来たため、雌雄の形態の違いやそれを作り出す仕組みは生物種によって様々ですが、性分化の究極の目的である生殖器官や生殖細胞を作る段階には、動物において一定の共通性が見出だされています。しかし植物の性分化においても、進化を通じて共通した制御因子が存在するのか、存在するとすればどのように働いているのかは、これまで全く分かっていませんでした。
中島・河内らの研究グループは、植物に共通した性分化の制御因子を見つけ出すため、陸上植物の進化系統樹で最も基部側に位置する苔類(たいるい)のゼニゴケ(Marchantia polymorpha)と、先端側に位置する被子植物のシロイヌナズナ(Arabidopsis thaliana)の比較解析を行いました(図1)。
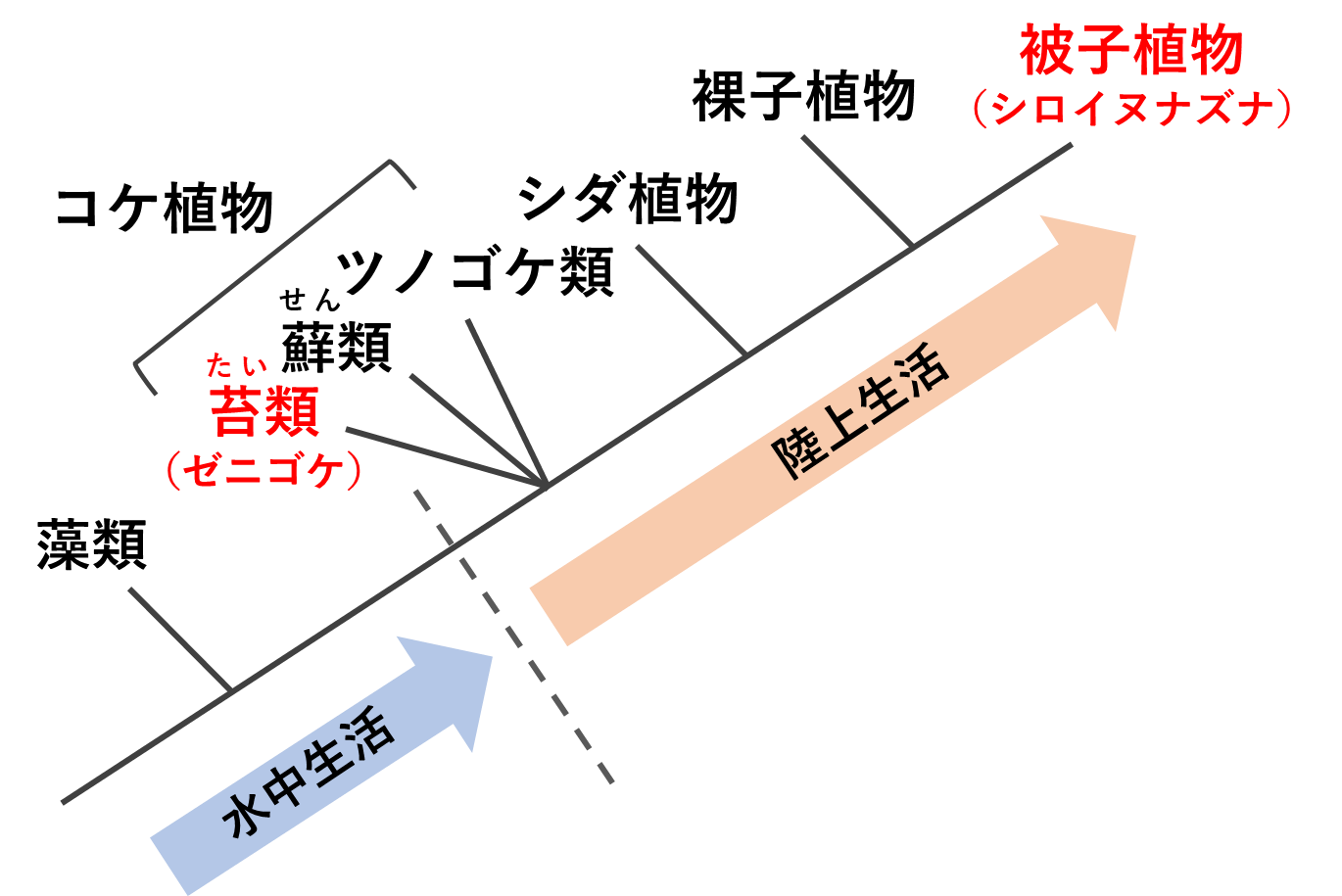
図1
ゼニゴケは河内らのグループが中心となって確立した我が国発のモデル植物であり、発生遺伝学研究に適した様々な利点を備えています。シロイヌナズナは世界的に汎用されているモデル被子植物です。この両者の比較により、最近になって陸上植物の進化に関する多くの重要な知見が得られるようになっています。
本研究においては両植物種の比較解析から、陸上植物に共通したメス分化の促進遺伝子FGMYBが発見され、さらにゼニゴケにおいては、この遺伝子の表裏の使いわけを用いた前例のない雌雄分化のスイッチング機構が明らかとなりました。
【実験の結果】
ゼニゴケ*1はメスとオスの個体が分かれた雌雄異株植物*2です。研究グループは、ゼニゴケのメスの生殖器官である造卵器(ぞうらんき)で強く発現する遺伝子を探索し、その結果を既に報告されているシロイヌナズナの胚嚢(はいのう)で発現する遺伝子のリストと照らし合わせました(図2)。胚嚢は、被子植物のめしべの中でメスの生殖機能の中心をなす部分であり、発生進化学的には生殖成長期のゼニゴケのメス個体全体に相当します*3。この比較解析からゼニゴケの造卵器と、シロイヌナズナの胚嚢で共通して強く発現している遺伝子を23組見出しました。これらのうちの1組はMYB型転写因子*4をコードする遺伝子でありFGMYBと名付けました(図2)。
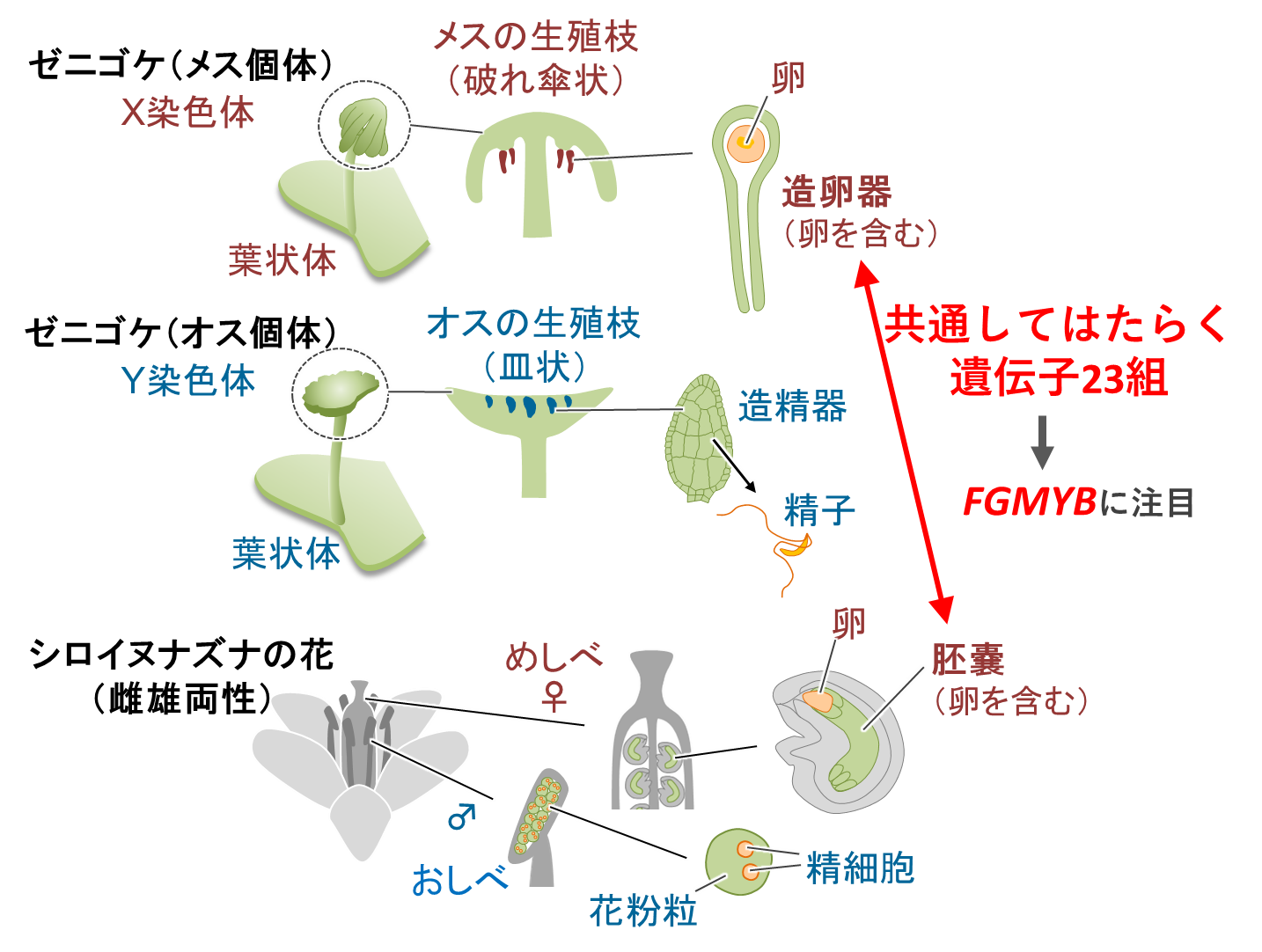
図2
ゼニゴケとシロイヌナズナの各々の遺伝子については、それぞれの学名の頭文字をとって、MpFGMYBあるいは AtFGMYBと呼びます。シロイヌナズナは転写因子をコードする遺伝子の数が多く、AtFGMYBについても3つの遺伝子がありました。これに対してゼニゴケのMpFGMYBは単一遺伝子でした。3つのAtFGMYB遺伝子については、過去に米国のグループが胚嚢の発達や機能に必要であることを報告しています。この報告をもとに、MpFGMYBも、ゼニゴケのメスの生殖機能に何らかの役割を果たしていると予測し、変異体を作って詳しく解析することにしました。
ゼニゴケはCRISPR/Cas9*5を用いたゲノム編集を比較的容易に行うことが出来ます。この方法を用いてMpFGMYBが機能しない株(Mpfgmyb変異体)をオス背景とメス背景の両方で作成しました。これらの変異体は、生殖成長期に入る前は野生株と同じ形態をしていました。
ところが生殖成長期に入ると、X染色体をもつメスであるはずの個体から、オスと見分けがつかない形態の生殖枝が形成されました。この生殖枝にはオスの生殖器官である造精器が作られ、その中には精子が作られました(図3)。ただし精子の運動性が野生型よりも悪く、受精能を失っていました。一方で、遺伝的にオスのMpfgmyb変異体は、外見も機能もオスのままでした(図3)。このことから、MpFGMYBはX染色体をもつ個体を正常なメスに分化させるために必須の遺伝子であることが分かりました。また、ゼニゴケの性分化の基底状態(デフォルト)はオスであり、メス個体でMpFGMYBがはたらかないと、基底状態であるオスの形態になることもわかりました。さらにオスとしての形態や精子形成に、Y染色体上の遺伝子はほぼ不要ですが、運動性を持つ精子を作るには、やはりY染色体上の遺伝子が関わっていることが明らかとなりました。
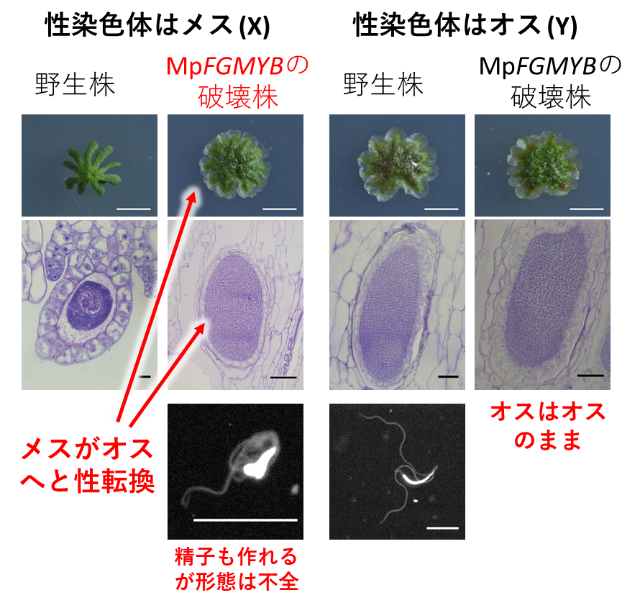
図3
MpFGMYB遺伝子はゼニゴケの常染色体上*6にあり、従って雌雄どちらのゲノムにも存在しています。しかしMpFGMYBは生殖成長期に入ったメスでのみはたらいていました。つまりオスにはMpFGMYB遺伝子のはたらきを抑える仕組みがあり、これがオスの性分化に重要であると考えられました。そこでゼニゴケのメスとオスの遺伝子発現を詳しく調べたところ、MpFGMYB遺伝子の裏側(DNA二本鎖の相補鎖側)から、オスのみで遺伝子発現が起こっていることがわかり、この裏側遺伝子*7をSUFと名付けました(図4)。
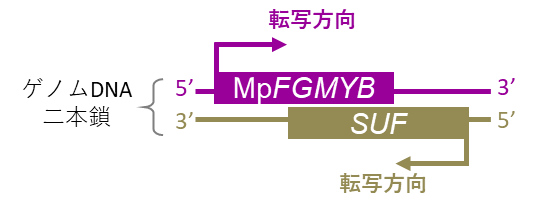
図4
SUFが機能しない変異体 (suf変異体)を作成したところ、Y染色体をもち、遺伝的にはオスであるはずの個体でもMpFGMYBが発現し、野生型のメス個体と区別できない形態を持つ生殖枝や造卵器が作られました(図5)。さらにMpFGMYBも破壊したところ、オスの形態に戻りました(図5)。

図5
以上の結果から、オス個体では裏側のSUFがはたらき、これが表側のMpFGMYBのはたらきを抑えていることが明らかとなりました(図6)。メス個体ではX染色体上のメス決定遺伝子によりSUFのはたらきが抑えられ、MpFGMYBの抑制が解かれてメスとしての形態が現れると考えられます(図6)。一方でsuf変異体のオスは、メスの生殖枝や造卵器を作るものの、卵細胞は作れませんでした。完全な卵を作るためには、X染色体上の遺伝子が必要であると考えられました(図5)。
一方で、上記のメス化したsuf変異体にSUF遺伝子を過剰発現する人工遺伝子を導入しても、suf変異体をオスに戻すことはできませんでした。またMpFGMYBとSUFを含む遺伝子断片をオスに導入してもオスのままでしたが、MpFGMYB遺伝子だけを導入するとメスへと性転換しました。これらの結果は、新たに導入したMpFGMYB遺伝子はゲノム上に元からあったSUF遺伝子により抑制されないものの、導入した同じ遺伝子断片上のSUFには抑制されることがわかりました。
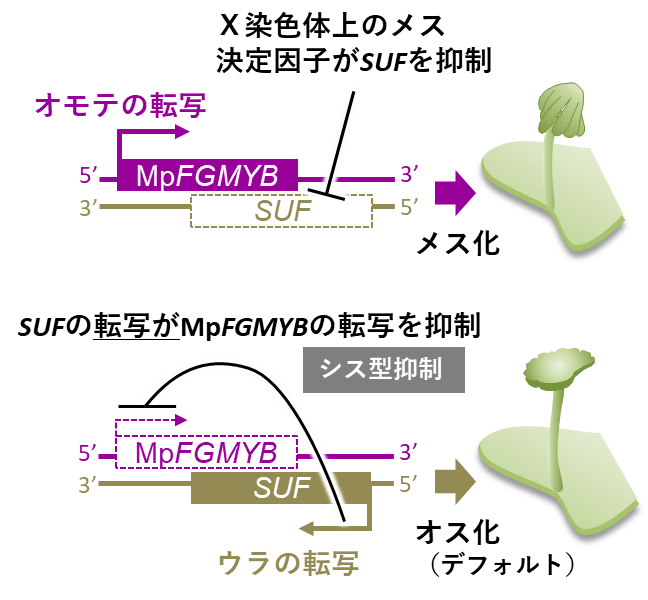
図6
つまりSUFがMpFGMYBを抑えられるのは、両者が同じ遺伝子の裏表に配置されている場合のみであり、これら2つの遺伝子が離れた場所にある場合には抑制できないことが分かりました。このことからSUFから作られたmRNA自体は抑制作用をもたず、SUF遺伝子の転写自体が、MpFGMYBのはたらきを抑えている(シスに働く)ことが分かりました(図7)。
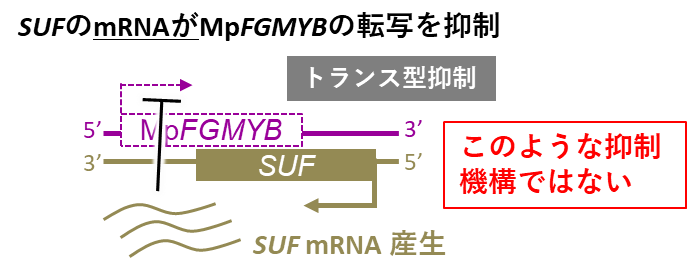
図7
以上の結果から、①ゼニゴケにおいてMpFGMYBがメスの性分化を制御するマスター因子であること、②MpFGMYBのはたらきが裏側のSUFにより直接抑制されることでオスの性分化がおこること、の2つが示されました。遺伝子の裏表を使った性分化のスイッチ機構はこれまでに報告がありません。他のコケ植物も同じスイッチ機構を用いているか、また他の生物でも同様の性分化機構があるのかは今のところ不明です。
またシロイヌナズナのAtFGMYB遺伝子の裏側にはSUFのような遺伝子は存在せず、AtFGMYBは、通常の遺伝子と同様に単純な転写制御を受けていると考えられます。複相世代が優勢となっている被子植物では、AtFGMYBのオン・オフが、複相のメス器官であるめしべの発達プログラムの下に組み込まれてしまっていると考えられます(図8)。
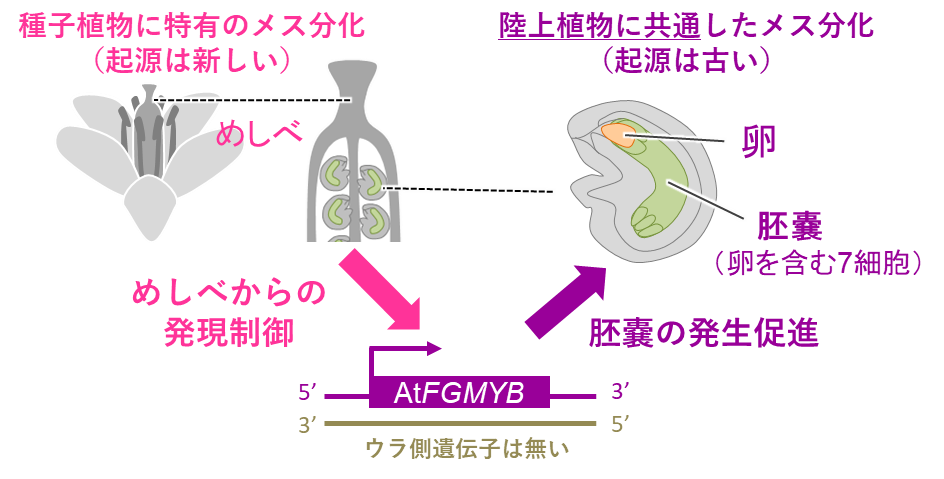
図8
【今後の発展】
本研究により、陸上植物の進化を通じて共通したメス分化の制御因子FGMYBが発見されました。FGMYBは、他の遺伝子のはたらきを調節する転写因子を作る遺伝子であるため、FGMYBを手掛かりとして、植物がメスとしての生殖能を発揮する際にはたらく遺伝子群を見つけ出せるようになります。ゼニゴケとシロイヌナズナは陸上植物の進化の両極にありますが、「卵細胞を作る」という有性生殖の本質にかかわる部分の制御には共通性を残していると考えられます。FGMYB遺伝子が制御する遺伝子をゼニゴケとシロイヌナズナで比較することで、植物のメス分化の何が共通で、何が変化しているのかを、学術的に明らかにすることが出来るようになります。このような知識は、種間雑種の作成や生殖能力の強化を目指した応用研究の基盤となるものであり、作物の効率的育種や繁殖能の改善につながります。
【用語解説】
- ゼニゴケ (liverwort)
陸上植物の進化系統樹の最も基部に位置するコケ植物苔類(たいるい)の1種。4億年以上前に初めて陸上生活を送るようになった共通祖先植物と似た形や性質を保持していると考えられている。転写因子の機能重複が少ないことや、形質転換やゲノム編集が容易であることなどから、コケ植物のモデルとして世界中で広く利用されている。シロイヌナズナやイネなどとの比較解析を通じ、陸上植物の進化の過程を明らかにする多くの研究成果が得られている。 - 雌雄異株(しゆういしゅ)と雌雄同株(しゆうどうしゅ)
同一個体に雌雄の性質を併せ持つ植物を雌雄同株、雌雄が別の個体に分かれた植物を雌雄異株と呼ぶ。これらの性質は植物の進化で何度も独立に現れており、その有無や態様は進化系統樹と一致しない。被子植物の多くは、両性花を咲かせる雌雄同株であるが、雄花や雌花といった単性花を別個体に咲かせる雌雄異株植物も良く知られている。ただし単性花は両性花のおしべ又はめしべが選択的に退化あるいは不稔化して生じたものであり、本研究で対象とした性分化とは、成り立ちが異なるものである(図9)。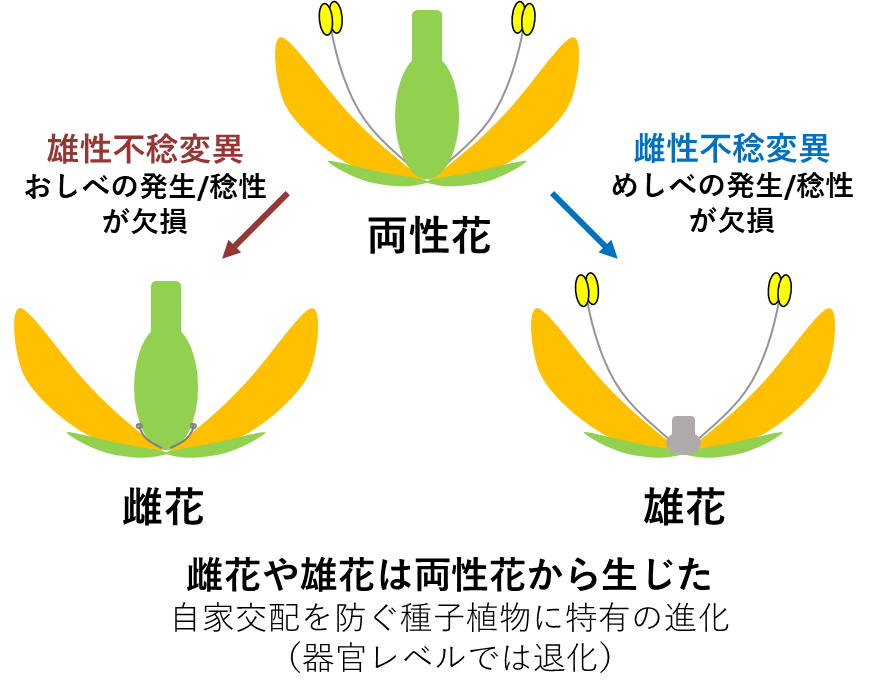
図9 - 陸上植物の生活環(図10)
植物の生活環は、減数分裂で作られた半数体(単相)細胞が作る配偶体と、半数体細胞が接合(受精)して作られる二倍体(複相)細胞が作る胞子体からなる。コケの生活環(図10左)は配偶体優勢であり、コケ個体は半数体(単相)である。栄養成長期のゼニゴケは雌雄の見分けがつかないが、生殖成長期になると性染色体で決定された性に従って異なる形の生殖枝が作られ、それらの中で卵や精子が成熟する。精子は泳いでメス個体内の卵と受精する。受精卵は数回の分裂により胞子嚢を作り、減数分裂を経て無数の胞子を作る。放出された胞子が発芽し、メスまたはオスの個体を作って栄養生殖期に戻るサイクルを繰り返す。
被子植物の生活環(図10右)は胞子体優勢であり、植物個体は二倍体(複相)の接合子から作られる。おしべやめしべという胞子体の性器官は植物進化の過程で比較的最近になって獲得されたものである。めしべの中で減数分裂がおこり、卵や中央細胞を含む7細胞からなる胚嚢が形成される。この胚嚢が被子植物の雌性配偶体であり、ゼニゴケのメス個体に相当する。おしべの中でも減数分裂がおこり、1つの栄養細胞が1つの雄原細胞を包み込んだ花粉粒となる。花粉粒が被子植物の雄性配偶体であり、ゼニゴケのオス個体に相当する。花粉粒の中で雄原細胞がさらに分裂して2つの精細胞となる。受粉により2つの精細胞の1つは卵と融合して受精卵となり胚を形成する。もう1つは中央細胞と融合して胚乳を形成する。
植物の生活環としては、現生コケ植物のような配偶体世代が優勢な方が祖先的である。被子植物に見られる胞子体世代が優勢な生活環は、比較的最近になって出現したものである。
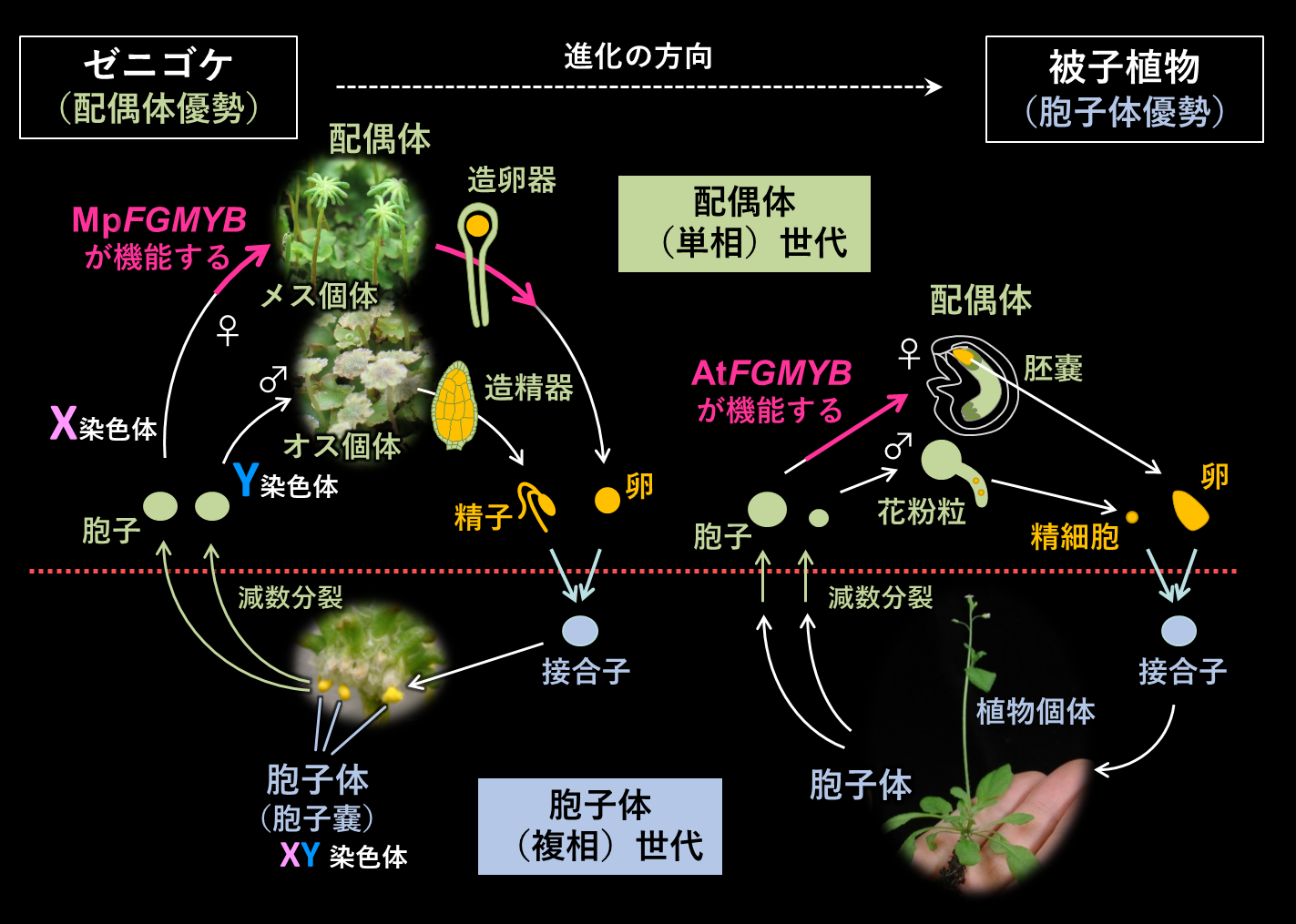
図10 - MYB型転写因子
ゲノムDNAの調節領域に結合し、mRNAへの転写を調節するタンパク質を転写因子と呼ぶ。転写を活性化する因子と抑制する因子がある。シロイヌナズナのゲノムには約2000個、ゼニゴケのゲノムには約400個の転写因子がコードされている。MYB型転写因子は真核生物に広く見られる転写因子ファミリーの1つであり、シロイヌナズナのゲノムには約200個、ゼニゴケのゲノムには約60個がコードされている。植物において、MYB型転写因子は様々な発生や生理現象に関わる遺伝子の転写を制御している。 - CRISPR/Cas9 (Clustered regularly interspaced short palindromic repeats (CRISPR)/CRISPR-associated 9)システム
細菌の獲得免疫システムにおいて機能するRNA依存型DNA切断酵素を用いたゲノム編集手法。2012年にCharpentierとDoudnaにより実用化が提唱されて以降、世界中で急速に普及し、様々な生物種のゲノム編集に利用されている。ゼニゴケでは、CRISPR/Cas9による効率的なゲノム編集技術が確立されている。 - 常染色体と性染色体
雌雄の性を決定する染色体を性染色体 (sex chromosome)、それ以外の染色体を常染色体(autosome)と呼ぶ。性染色体には性を決定する遺伝子が乗っているが、各性に特有の形態や生殖機能(性分化)を司る遺伝子がすべて性染色体に乗っているわけではない。むしろ性染色体には性決定遺伝子だけが存在し、その遺伝子産物が常染色体上の性分化関連遺伝子を調節することで性分化を誘導している例も多い。 - 遺伝子の表と裏
遺伝子の「表と裏」は比喩的な表現であり、専門的には「センスとアンチセンス」と表現される。遺伝子の実体はDNA二本鎖である。一般に「遺伝子が発現する」とは、DNA二本鎖のうち特定の一本(アンチセンス鎖)を鋳型としてその配列情報をメッセンジャーRNA (mRNA)に写し取り(転写し)、その配列情報をアミノ酸配列へと「翻訳」してタンパク質を作ることを言う。mRNAは、DNAのアンチセンス鎖を鋳型として作られるため、センス鎖の配列を持つ(図11)。もしDNAのセンス鎖を鋳型としてmRNAが合成されても、できたmRNAはアンチセンス鎖の配列をもつため正しいアミノ酸配列へ翻訳されず機能しない。したがって、DNA二本鎖のうちのどちらがmRNAに転写されるかは、遺伝子ごとに厳密に決められている(図12)。MpFGMYBとSUFのように、両方のDNA鎖から機能的なmRNAが作られる遺伝子は非常にまれであるが、動植物を通じていくつかの報告例がある。
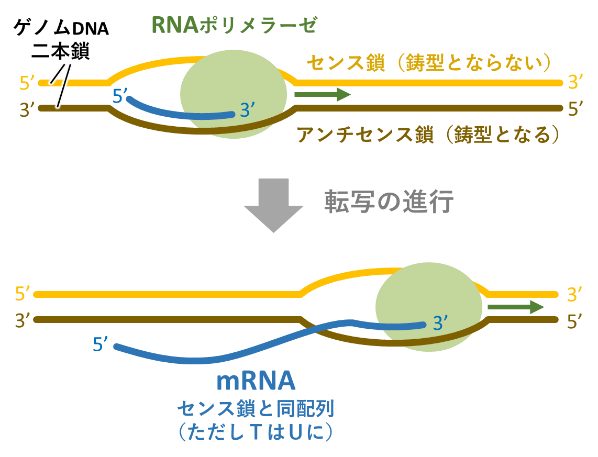
図11
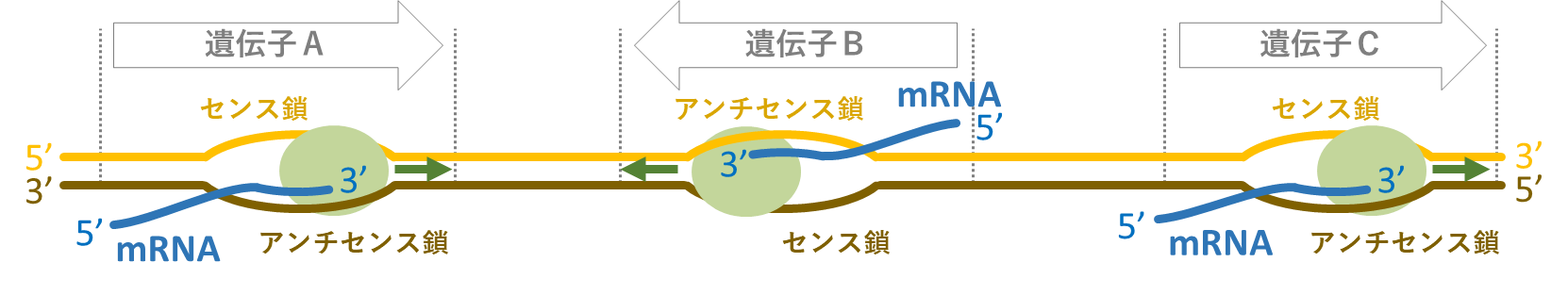
図12
【本プレスリリースに関するお問い合わせ先】
奈良先端科学技術大学院大学 先端科学技術研究科
バイオサイエンス領域 植物発生シグナル研究室 中島 敬二(なかじま けいじ)教授
E-mail: k-nakaji@bs.naist.jp